

A glycine-rich RNA-binding protein mediating cold-inducible suppression of mammalian cell growth
- PMID: 9151692
- PMCID: PMC2139845
- DOI: 10.1083/jcb.137.4.899
A glycine-rich RNA-binding protein mediating cold-inducible suppression of mammalian cell growth
Abstract
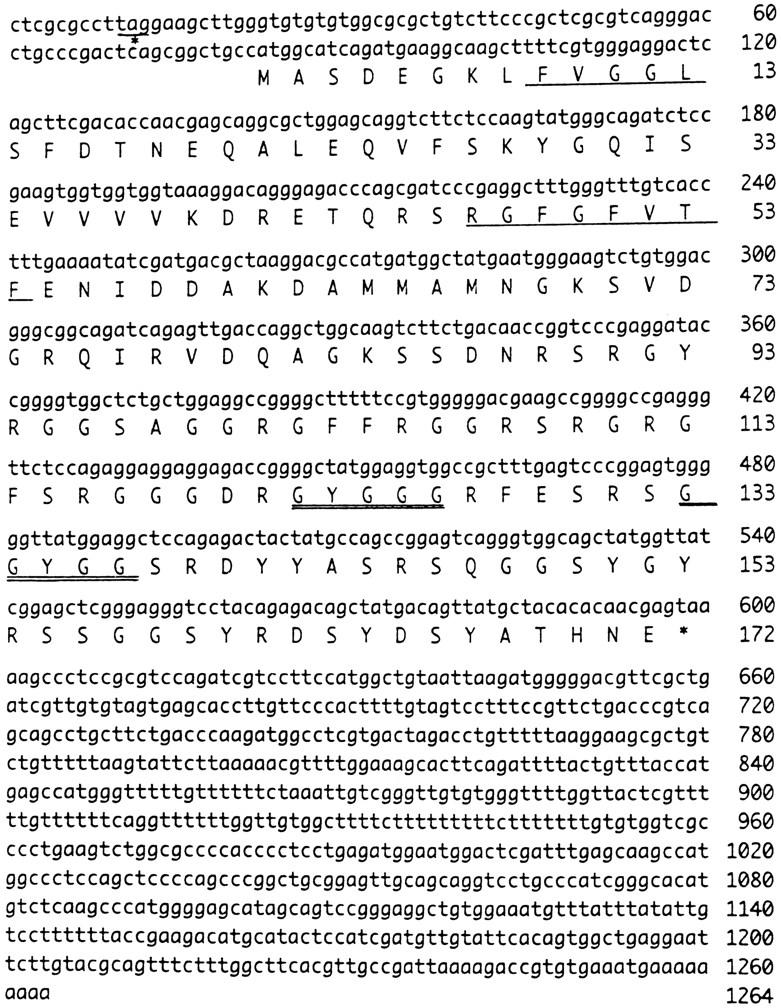
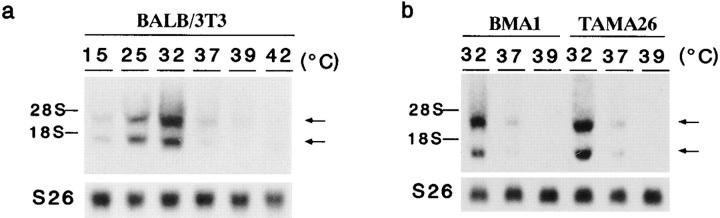
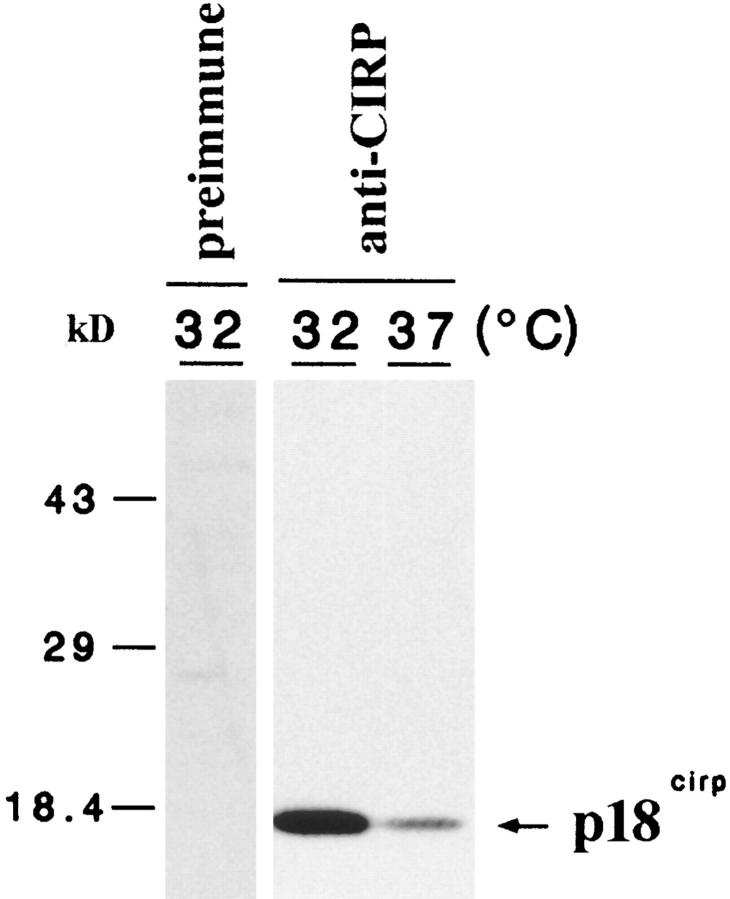
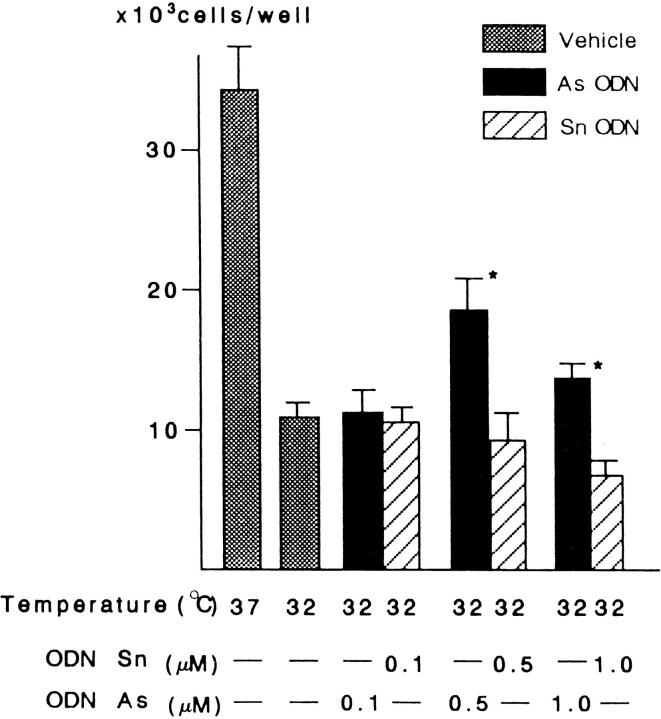
In response to low ambient temperature, mammalian cells as well as microorganisms change various physiological functions, but the molecular mechanisms underlying these adaptations are just beginning to be understood. We report here the isolation of a mouse cold-inducible RNA-binding protein (cirp) cDNA and investigation of its role in cold-stress response of mammalian cells. The cirp cDNA encoded an 18-kD protein consisting of an amino-terminal RNAbinding domain and a carboxyl-terminal glycine-rich domain and exhibited structural similarity to a class of stress-induced RNA-binding proteins found in plants. Immunofluorescence microscopy showed that CIRP was localized in the nucleoplasm of BALB/3T3 mouse fibroblasts. When the culture temperature was lowered from 37 to 32 degrees C, expression of CIRP was induced and growth of BALB/3T3 cells was impaired as compared with that at 37 degrees C. By suppressing the induction of CIRP with antisense oligodeoxynucleotides, this impairment was alleviated, while overexpression of CIRP resulted in impaired growth at 37 degrees C with prolongation of G1 phase of the cell cycle. These results indicate that CIRP plays an essential role in cold-induced growth suppression of mouse fibroblasts. Identification of CIRP may provide a clue to the regulatory mechanisms of cold responses in mammalian cells.
Figures







References
-
- Aloia RC, Raison JK. Membrane function in mammalian hibernation. Biochim Biophys Acta. 1989;988:123–146. - PubMed
-
- Bandziulis RJ, Swanson MS, Dreyfuss G. RNA-binding proteins as developmental regulators. Genes Dev. 1989;3:431–437. - PubMed
-
- Bergeron D, Beauseigle D, Bellemare G. Sequence and expression of a gene encoding a protein with RNA-binding and glycine-rich domains in Brassica napus. . Biochim Biophys Acta. 1993;1216:123–125. - PubMed
-
- Burd CG, Dreyfuss G. Conserved structures and diversity of functions of RNA-binding proteins. Science (Wash DC) 1994;265:615–621. - PubMed
-
- Burdon RH. Temperature and animal cell protein synthesis. Symp Soc Exp Biol. 1987;41:113–133. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
