

Neurosteroid prolongs GABAA channel deactivation by altering kinetics of desensitized states
- PMID: 9151718
- PMCID: PMC6573570
- DOI: 10.1523/JNEUROSCI.17-11-04022.1997
Neurosteroid prolongs GABAA channel deactivation by altering kinetics of desensitized states
Abstract
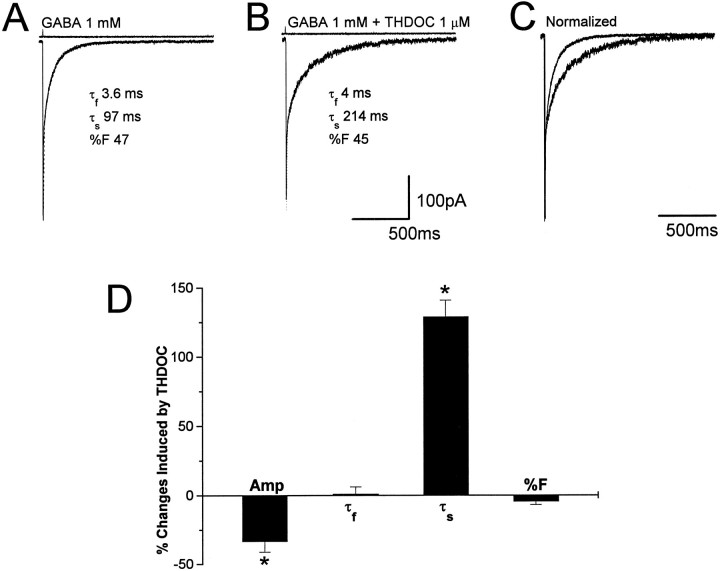
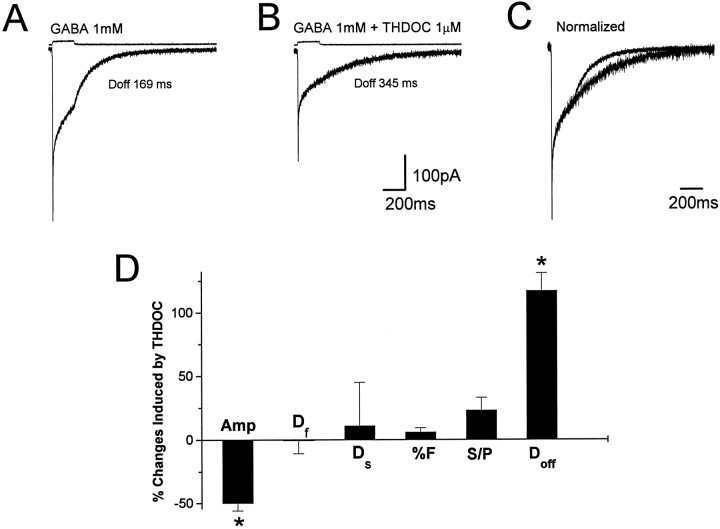
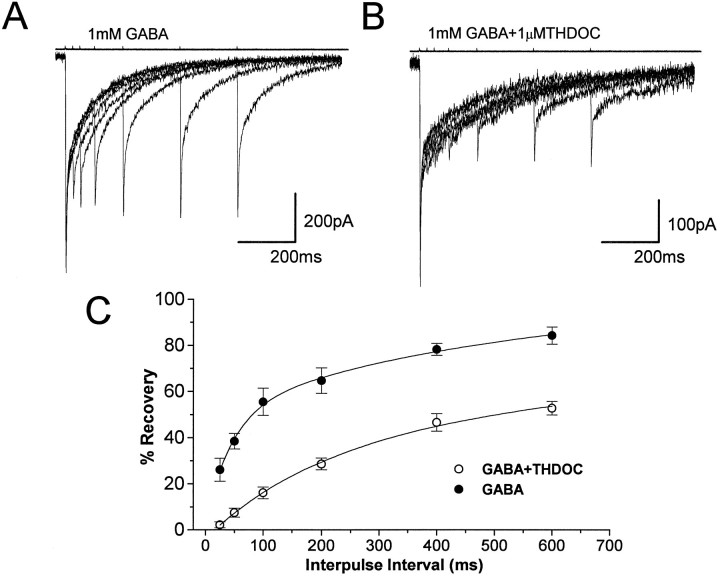
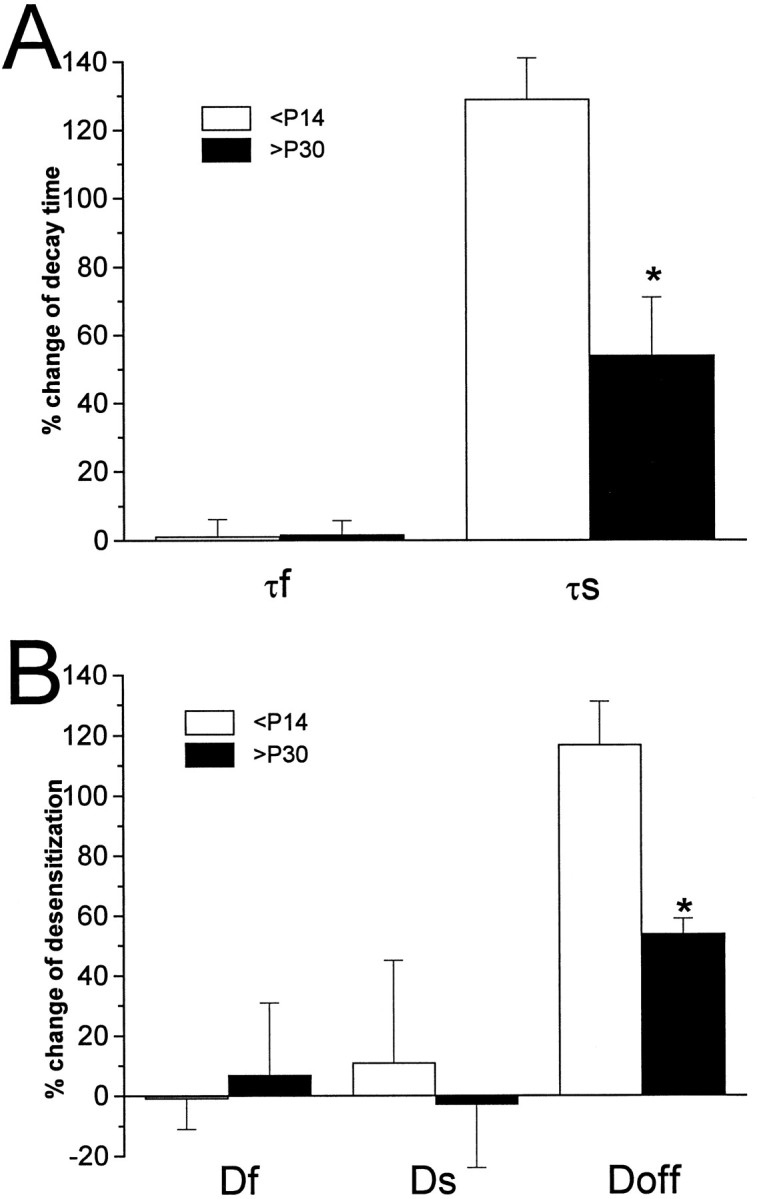
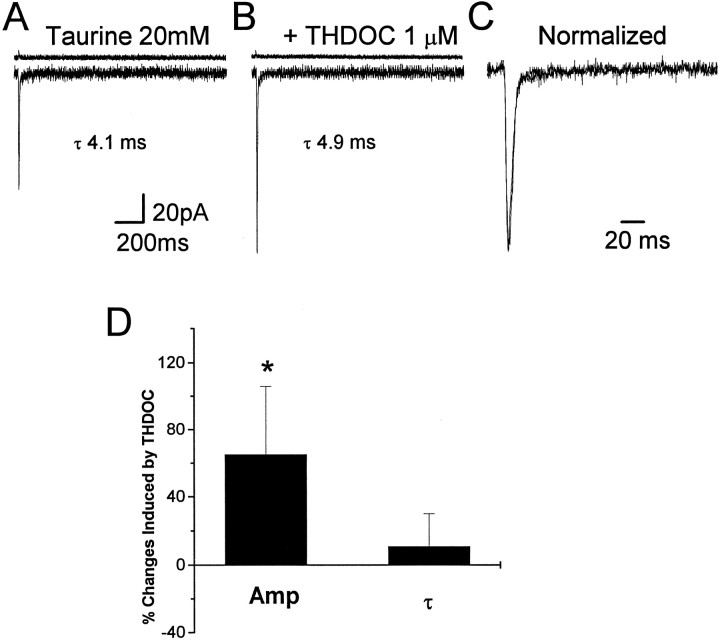
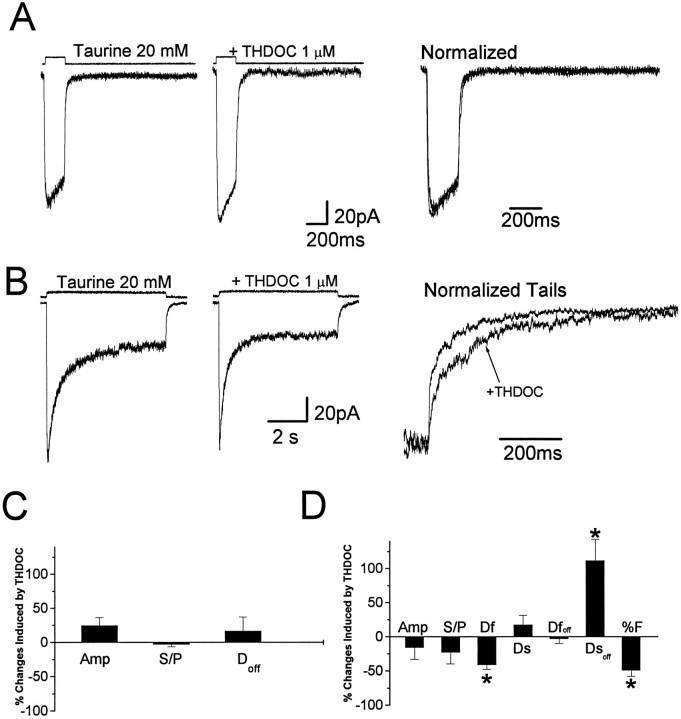
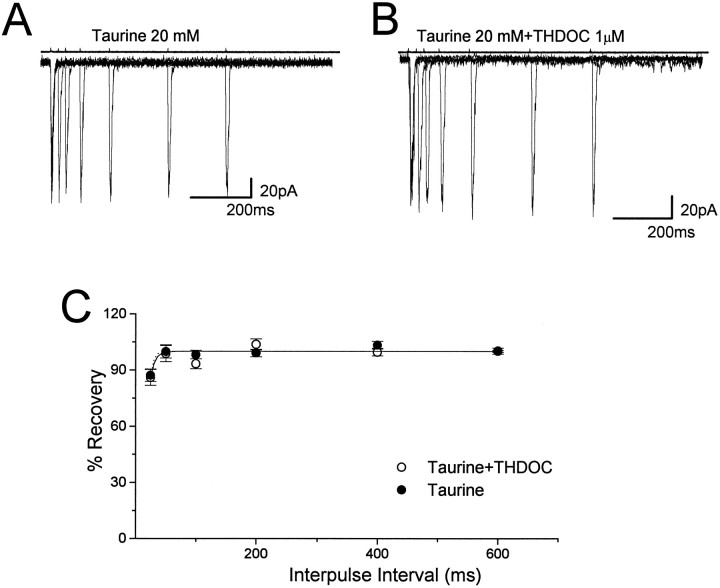
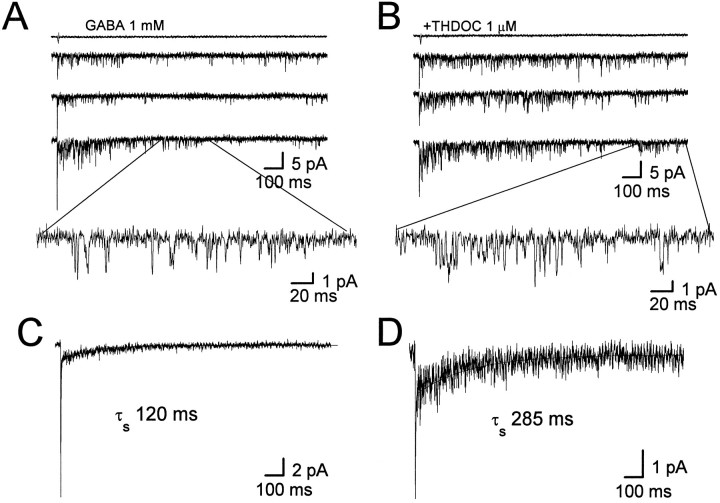
Fast applications of GABA (1 mM) to nucleated and outside-out patches excised from granule neurons in cerebellar slices from developing rats evoked currents with a double exponential time course reminiscent of that of IPSCs. A neurosteroid 3alpha, 21dihydroxy-5alpha-pregnan-20-one (THDOC) remarkably increased the slow deactivation time constant and slowed down recovery from desensitization, as estimated by paired-pulse GABA applications. THDOC also reduced the amplitude of GABA currents, whereas it failed to affect the fast deactivation component and its relative contribution to peak amplitude. The effects of THDOC on slow deactivation were greater in rats younger than postnatal day 13 (P13) as compared with rats at P30-P35. THDOC failed to alter deactivation of short responses induced by a less-potent agonist taurine at saturating doses. These responses had deactivation kinetics described by a fast single exponential decay, little desensitization, and quick recovery. However, THDOC slowed deactivation if taurine responses were long enough to allow consistent desensitization, suggesting that desensitized states are required for the neurosteroid to modulate GABA responses. In outside-out patches, just as desensitized states prolonged GABA responses by producing reopening of channels activated by brief GABA pulses, THDOC increased the channel open probability by further increasing the number of late channel openings, resulting in a prolongation of the slow deactivation. Our data suggest that neurosteroid potentiates the inhibitory postsynaptic transmission via the prolongation of the slow deactivation and that the alteration of kinetics of entry and exit from desensitized states underlies the allosteric modification of GABAA receptors by neurosteroids.
Figures
References
-
- Callachan H, Cottrell GA, Hather NY, Lambert JJ, Nooney JM, Peters JA. Modulation of GABAA receptor by progesterone metabolites. Proc R Soc Lond [Biol] 1987;231:359–369. - PubMed
-
- Cooper EJ, Johnston GAR, Edwards FA. Differential sensitivity of synaptic GABAergic currents to a neuroactive steroid in brain slices from male rats. Soc Neurosci Abstr. 1995;21:1345.
-
- Cooper EJ, Johnston GAR, Edwards FA. Developmental differences in synaptic GABAergic currents in hippocampal and cerebellar cells of male rats. Soc Neurosci Abstr. 1996;22:810.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials