

Molecular analysis of the putative inactivation particle in the inactivation gate of brain type IIA Na+ channels
- PMID: 9154906
- PMCID: PMC2217064
- DOI: 10.1085/jgp.109.5.589
Molecular analysis of the putative inactivation particle in the inactivation gate of brain type IIA Na+ channels
Abstract
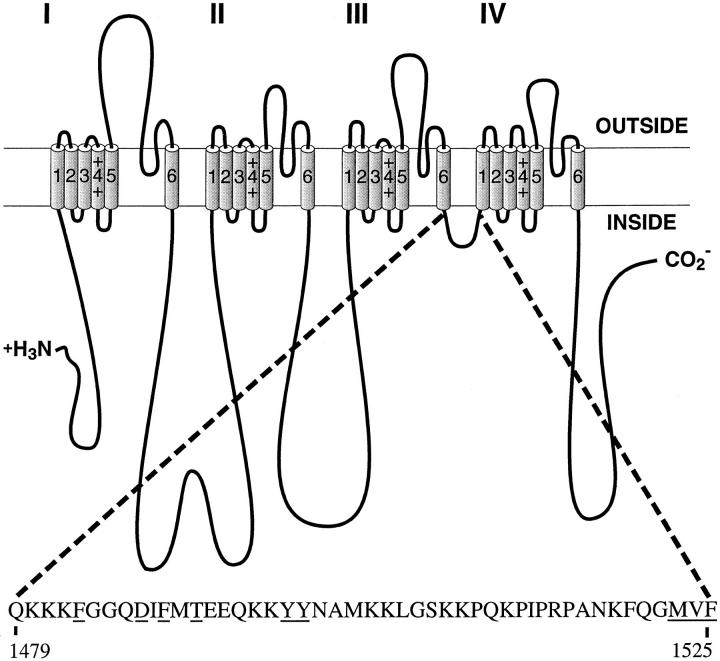
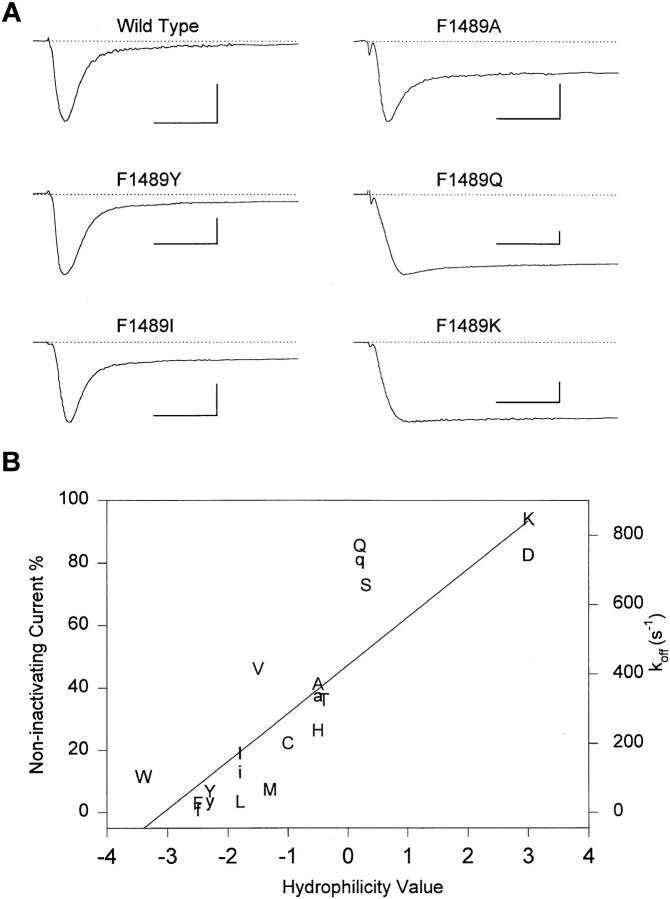
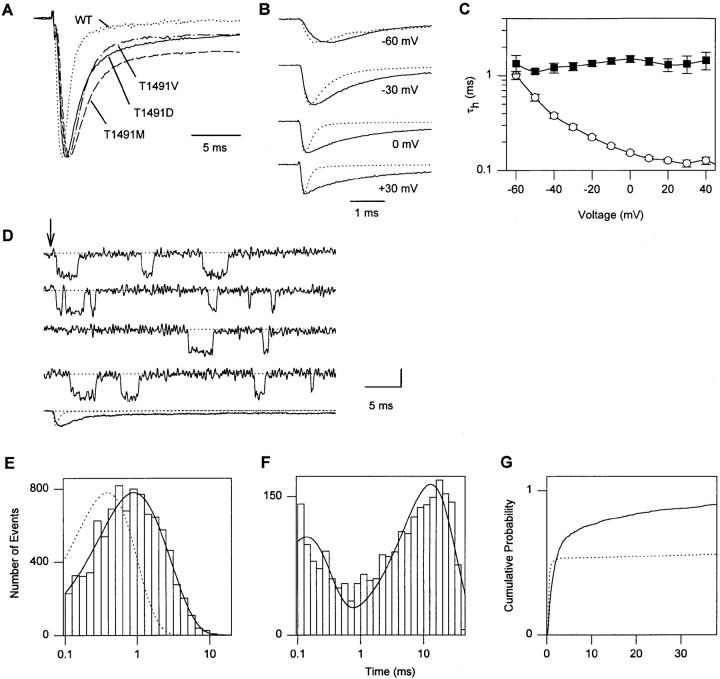
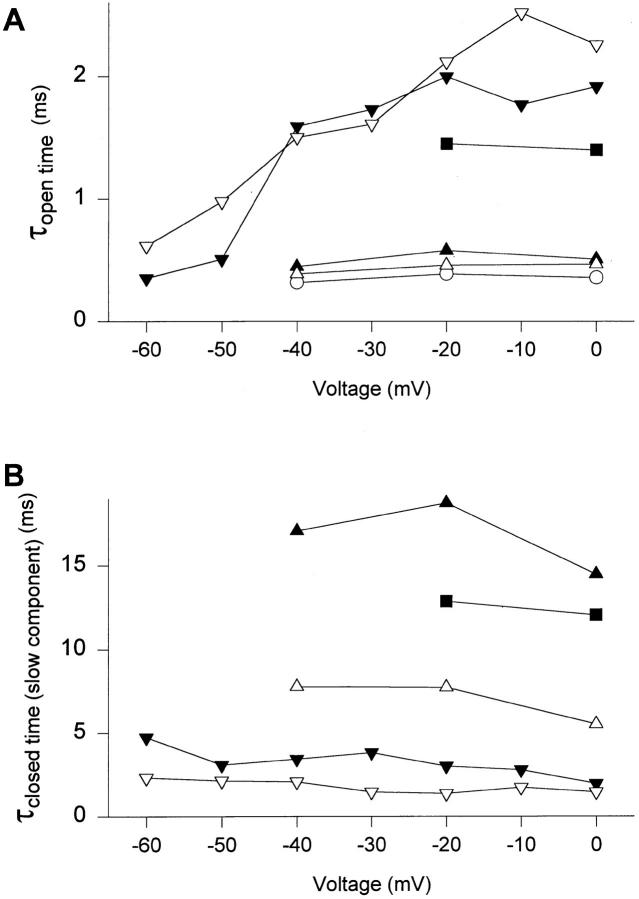
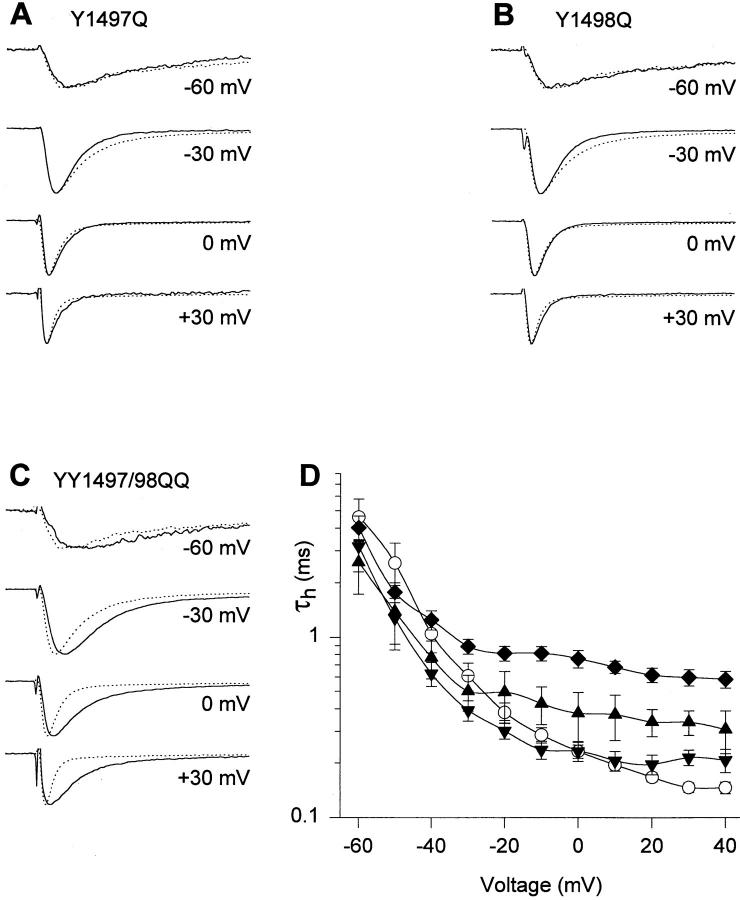
Fast Na+ channel inactivation is thought to involve binding of phenylalanine 1489 in the hydrophobic cluster IFM in L(III-IV) of the rat brain type IIA Na+ channel. We have analyzed macroscopic and single channel currents from Na+ channels with mutations within and adjacent to hydrophobic clusters in L(III-IV). Substitution of F1489 by a series of amino acids disrupted inactivation to different extents. The degree of disruption was closely correlated with the hydrophilicity of the amino acid at position 1489. These mutations dramatically destabilized the inactivated state and also significantly slowed the entry into the inactivated state, consistent with the idea that F1489 forms a hydrophobic interaction with a putative receptor during the fast inactivation process. Substitution of a phe residue at position 1488 or 1490 in mutants lacking F1489 did not restore normal inactivation, indicating that precise location of F1489 is critical for its function. Mutations of T1491 disrupted inactivation substantially, with large effects on the stability of the inactivated state and smaller effects on the rate of entry into the inactivated state. Mutations of several other hydrophobic residues did not destabilize the inactivated state at depolarized potentials, indicating that the effects of mutations at F1489 and T1491 are specific. The double mutant YY1497/8QQ slowed macroscopic inactivation at all potentials and accelerated recovery from inactivation at negative membrane potentials. Some of these mutations in L(III-IV) also affected the latency to first opening, indicating coupling between L(III-IV) and channel activation. Our results show that the amino acid residues of the IFM hydrophobic cluster and the adjacent T1491 are unique in contributing to the stability of the inactivated state, consistent with the designation of these residues as components of the inactivation particle responsible for fast inactivation of Na+ channels.
Figures





Similar articles
-
A critical role for transmembrane segment IVS6 of the sodium channel alpha subunit in fast inactivation.J Biol Chem. 1995 May 19;270(20):12025-34. doi: 10.1074/jbc.270.20.12025. J Biol Chem. 1995. PMID: 7744852
-
Molecular analysis of potential hinge residues in the inactivation gate of brain type IIA Na+ channels.J Gen Physiol. 1997 May;109(5):607-17. doi: 10.1085/jgp.109.5.607. J Gen Physiol. 1997. PMID: 9154907 Free PMC article.
-
A critical role for the S4-S5 intracellular loop in domain IV of the sodium channel alpha-subunit in fast inactivation.J Biol Chem. 1998 Jan 9;273(2):1121-9. doi: 10.1074/jbc.273.2.1121. J Biol Chem. 1998. PMID: 9422778
-
[Structures of peptides related to the inactivation gate on sodium channels].Yakugaku Zasshi. 2002 Dec;122(12):1123-31. doi: 10.1248/yakushi.122.1123. Yakugaku Zasshi. 2002. PMID: 12510389 Review. Japanese.
-
Molecular properties of brain sodium channels: an important target for anticonvulsant drugs.Adv Neurol. 1999;79:441-56. Adv Neurol. 1999. PMID: 10514834 Review.
Cited by
-
Action potential energy efficiency varies among neuron types in vertebrates and invertebrates.PLoS Comput Biol. 2010 Jul 1;6(7):e1000840. doi: 10.1371/journal.pcbi.1000840. PLoS Comput Biol. 2010. PMID: 20617202 Free PMC article.
-
A Mechanistic Reinterpretation of Fast Inactivation in Voltage-Gated Na + Channels.bioRxiv [Preprint]. 2023 Apr 28:2023.04.27.538555. doi: 10.1101/2023.04.27.538555. bioRxiv. 2023. Update in: Nat Commun. 2023 Aug 21;14(1):5072. doi: 10.1038/s41467-023-40514-4. PMID: 37162849 Free PMC article. Updated. Preprint.
-
Voltage-Gated Na+ Channels: Not Just for Conduction.Cold Spring Harb Perspect Biol. 2016 Jun 1;8(6):a029264. doi: 10.1101/cshperspect.a029264. Cold Spring Harb Perspect Biol. 2016. PMID: 27252364 Free PMC article. Review.
-
Mechanisms of closed-state inactivation in voltage-gated ion channels.J Physiol. 2011 Feb 1;589(Pt 3):461-79. doi: 10.1113/jphysiol.2010.191965. Epub 2010 Nov 22. J Physiol. 2011. PMID: 21098008 Free PMC article. Review.
-
Inactivation and recovery of sodium currents in cerebellar Purkinje neurons: evidence for two mechanisms.Biophys J. 2001 Feb;80(2):729-37. doi: 10.1016/S0006-3495(01)76052-3. Biophys J. 2001. PMID: 11159440 Free PMC article.
