

Reactivation of silenced, virally transduced genes by inhibitors of histone deacetylase
- PMID: 9159154
- PMCID: PMC20860
- DOI: 10.1073/pnas.94.11.5798
Reactivation of silenced, virally transduced genes by inhibitors of histone deacetylase
Abstract
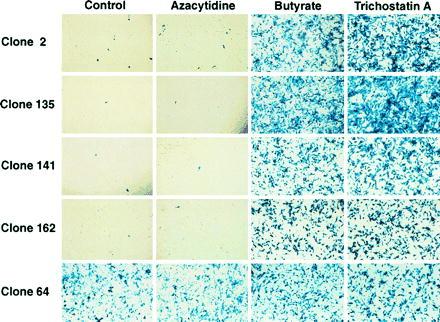
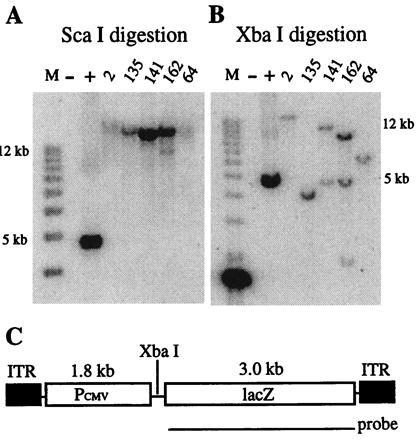
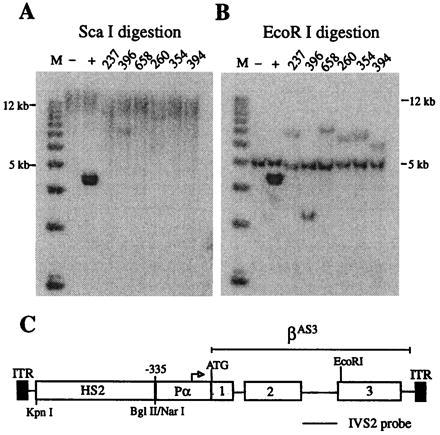
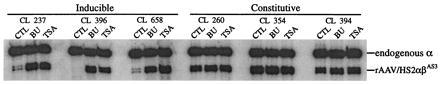
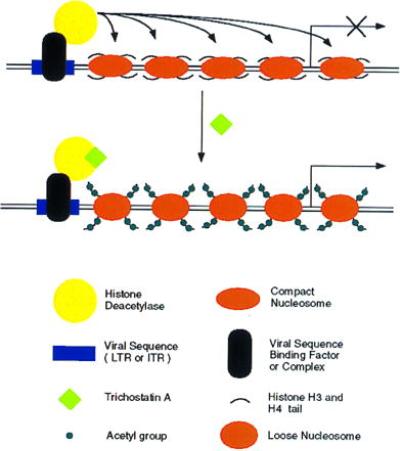
Retroviral and adeno-associated viral sequences can dramatically silence transgene expression in mice. We now report that this repression also occurs in stably infected HeLa cells when the cells are grown without selection. Expression of a transduced lacZ gene (rAAV/CMVlacZ) is silenced in greater than 90% of cells after 60 days in culture. Surprisingly, high-level expression can be reactivated by treating the cells with sodium butyrate or trichostatin A but not with 5-azacytidine. When cell clones with integrated copies of rAAV/CMVlacZ were isolated, lacZ expression was silenced in 80% of the clones; however, lacZ expression was reactivated in all of the silenced clones by treatment with butyrate or trichostatin A. The two drugs also reactivated a silenced globin gene construct (rAAV/HS2alphabetaAS3) in stably infected K562 cells. Trichostatin A is a specific inhibitor of histone deacetylase; therefore, we propose that hyperacetylation of histones after drug treatment changes the structure of chromatin on integrated viral sequences and relieves repression of transduced genes. The reactivation of silenced, transduced genes has implications for gene therapy. Efficient viral gene transfer followed by drug treatment to relieve suppression may provide a powerful combination for treatment of various genetic and infectious diseases.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
