

Neurotoxicity associated with dual actions of homocysteine at the N-methyl-D-aspartate receptor
- PMID: 9159176
- PMCID: PMC20882
- DOI: 10.1073/pnas.94.11.5923
Neurotoxicity associated with dual actions of homocysteine at the N-methyl-D-aspartate receptor
Abstract
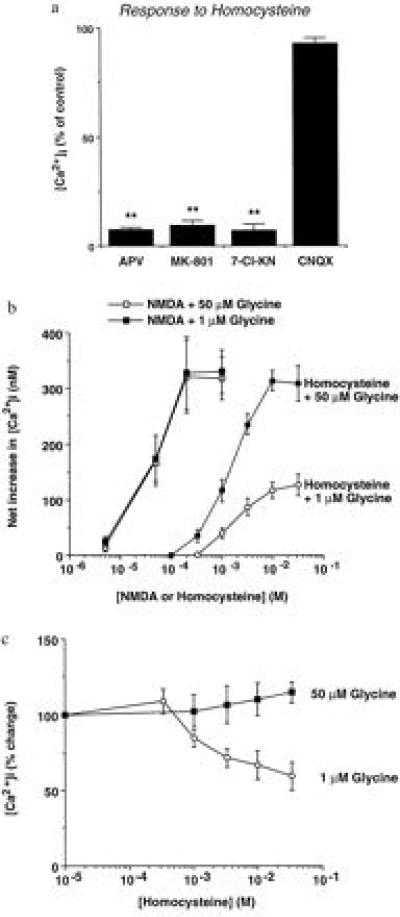
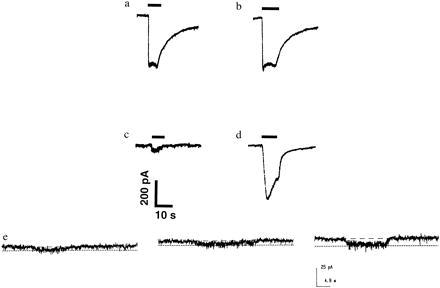
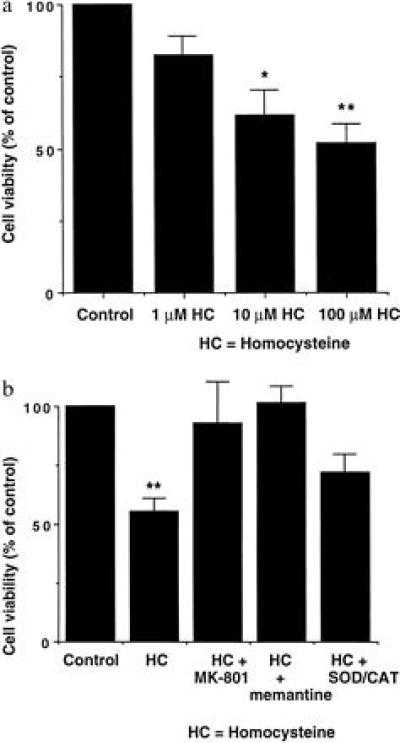
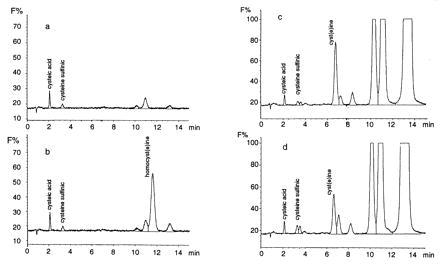
Severely elevated levels of total homocysteine (approximately millimolar) in the blood typify the childhood disease homocystinuria, whereas modest levels (tens of micromolar) are commonly found in adults who are at increased risk for vascular disease and stroke. Activation of the coagulation system and adverse effects of homocysteine on the endothelium and vessel wall are believed to underlie disease pathogenesis. Here we show that homocysteine acts as an agonist at the glutamate binding site of the N-methyl-D-aspartate receptor, but also as a partial antagonist of the glycine coagonist site. With physiological levels of glycine, neurotoxic concentrations of homocysteine are on the order of millimolar. However, under pathological conditions in which glycine levels in the nervous system are elevated, such as stroke and head trauma, homocysteine's neurotoxic (agonist) attributes at 10-100 microM levels outweigh its neuroprotective (antagonist) activity. Under these conditions neuronal damage derives from excessive Ca2+ influx and reactive oxygen generation. Accordingly, homocysteine neurotoxicity through overstimulation of N-methyl-D-aspartate receptors may contribute to the pathogenesis of both homocystinuria and modest hyperhomocysteinemia.
Figures
References
-
- Lindgren A, Brattström L, Norrving B, Hultberg B, Anderson A, Johannson B B. Stroke. 1995;26:795–800. - PubMed
-
- Perry I J, Refsum H, Morris R W, Ebrahim S B, Ueland P M, Shaper A G. Lancet. 1995;346:1395–1398. - PubMed
-
- Broch O J, Ueland P M. J Neurochem. 1984;43:1755–1757. - PubMed
-
- Stampfer M J, Malinow M R. N Engl J Med. 1995;332:328–329. - PubMed
-
- Perry T L, Bergeron C, Steele J C, McLachlan D R, Hansen S. J Neurol Sci. 1990;99:3–8. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
