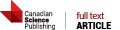The importance of the microenvironment in breast cancer progression: recapitulation of mammary tumorigenesis using a unique human mammary epithelial cell model and a three-dimensional culture assay
- PMID: 9164652
- PMCID: PMC2933195
- DOI: 10.1139/o96-089
The importance of the microenvironment in breast cancer progression: recapitulation of mammary tumorigenesis using a unique human mammary epithelial cell model and a three-dimensional culture assay
Abstract
The extracellular matrix (ECM) is a dominant regulator of tissue development and homeostasis. "Designer microenvironments" in culture and in vivo model systems have shown that the ECM regulates growth, differentiation, and apoptosis in murine and human mammary epithelial cells (MEC) through a hierarchy of transcriptional events involving the intricate interplay between soluble and physical signaling pathways. Furthermore, these studies have shown that these pathways direct and in turn are influenced by the tissue structure. Tissue structure is directed by the cooperative interactions of the cell-cell and cell-ECM pathways and can be modified by stromal factors. Not surprisingly then, loss of tissue structure and alterations in ECM components are associated with the appearance and dissemination of breast tumors, and malignancy is associated with perturbations in cell adhesion, changes in adhesion molecules, and a stromal reaction. Several lines of evidence now support the contention that the pathogenesis of breast cancer is determined (at least in part) by the dynamic interplay between the ductal epithelial cells, the microenvironment, and the tissue structure (acini). Thus, to understand the mechanisms involved in carcinogenesis, the role of the microenvironment (ECM as well as the stromal cells) with respect to tissue structure should be considered and studied. Towards this goal, we have established a unique human MEC model of tumorigenesis, which in concert with a three-dimensional assay, recapitulates many of the genetic and morphological changes observed in breast in cancer in vivo. We are currently using this system to understand the role of the microenvironment and tissue structure in breast cancer progression.
Figures





References
-
- Adams JC, Watt FM. Regulation of development and differentiation by the extracellular matrix. Development. 1993;117:1183–1198. - PubMed
-
- Alford D, Taylor-Papadimitriou J. Cell adhesion molecules in the normal and cancerous mammary gland. J. Mammary Gland Biol. Neoplasms. 1996;1:207–218. - PubMed
-
- Band V. Preneoplastic transformation of human mammary epithelial cells. Semin. Cell Biol. 1995;6:185–192. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical