

Site-specific and sensory neuron-dependent increases in postsynaptic glutamate sensitivity accompany serotonin-induced long-term facilitation at Aplysia sensorimotor synapses
- PMID: 9185535
- PMCID: PMC6573320
- DOI: 10.1523/JNEUROSCI.17-13-04976.1997
Site-specific and sensory neuron-dependent increases in postsynaptic glutamate sensitivity accompany serotonin-induced long-term facilitation at Aplysia sensorimotor synapses
Abstract
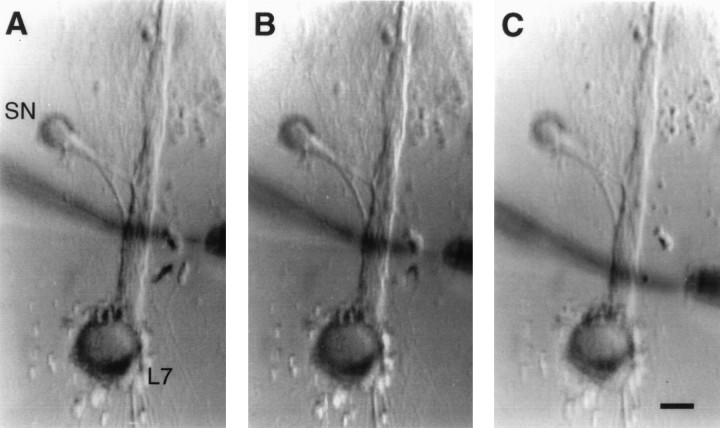
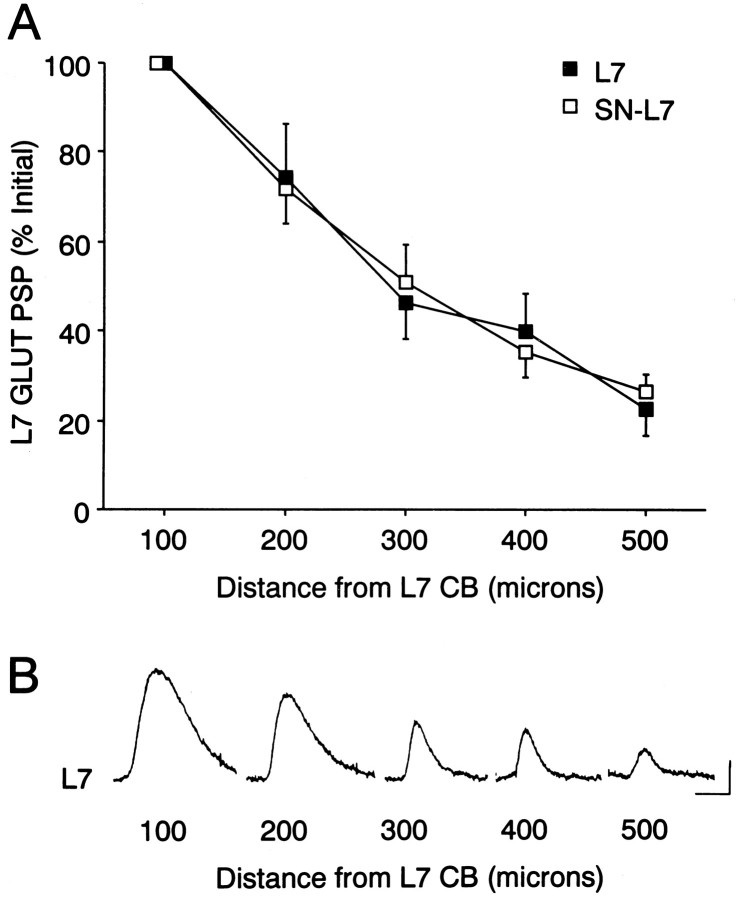
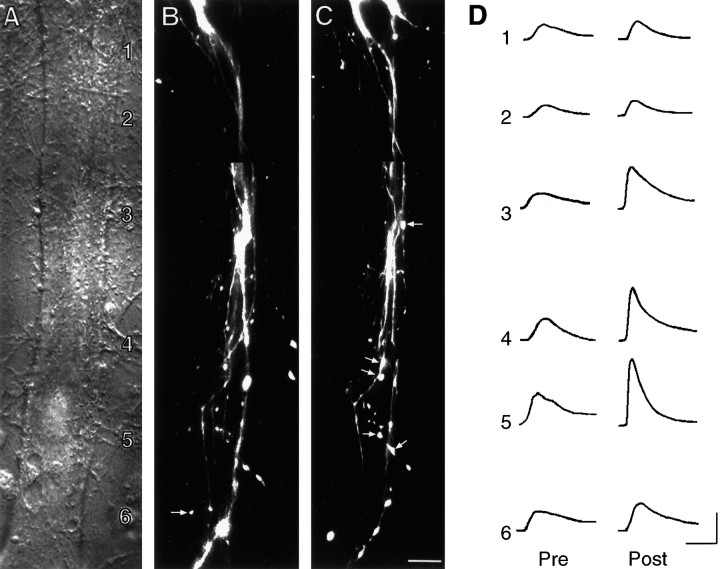
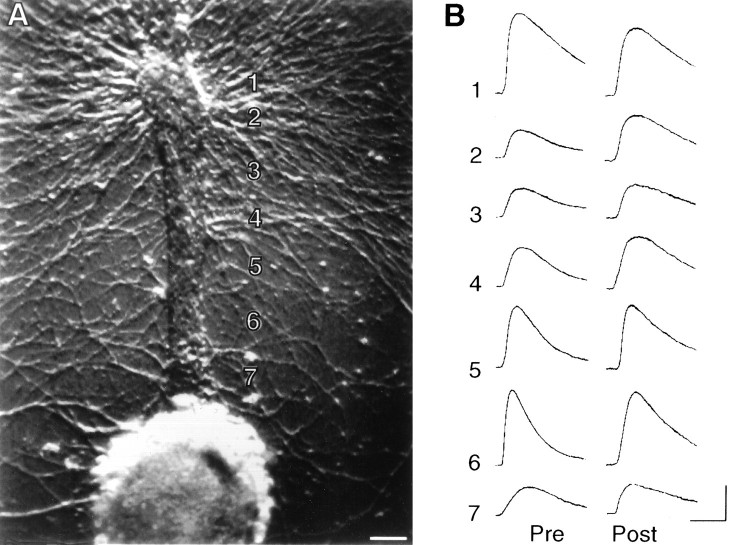
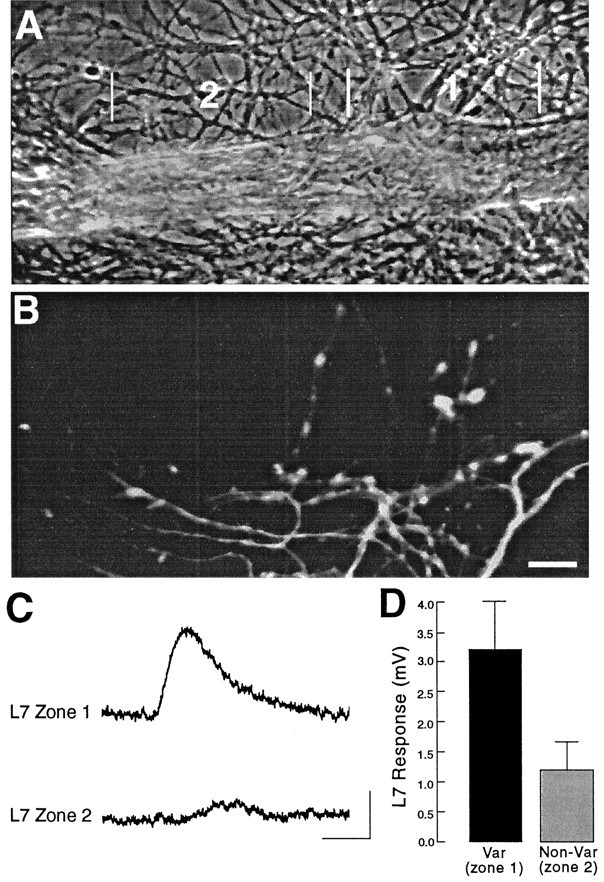
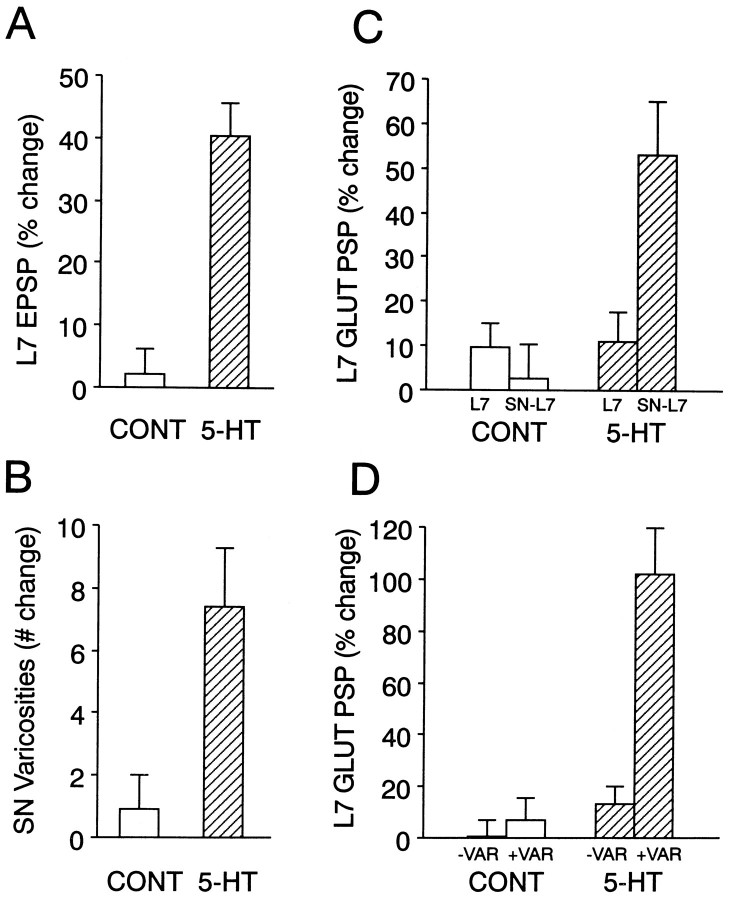
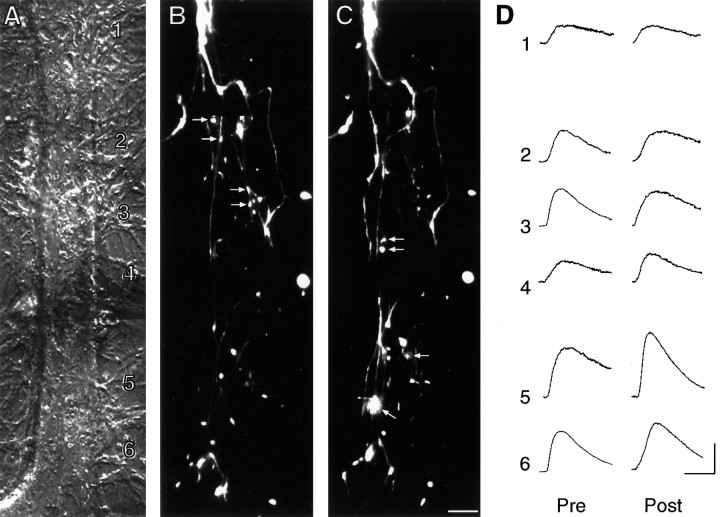
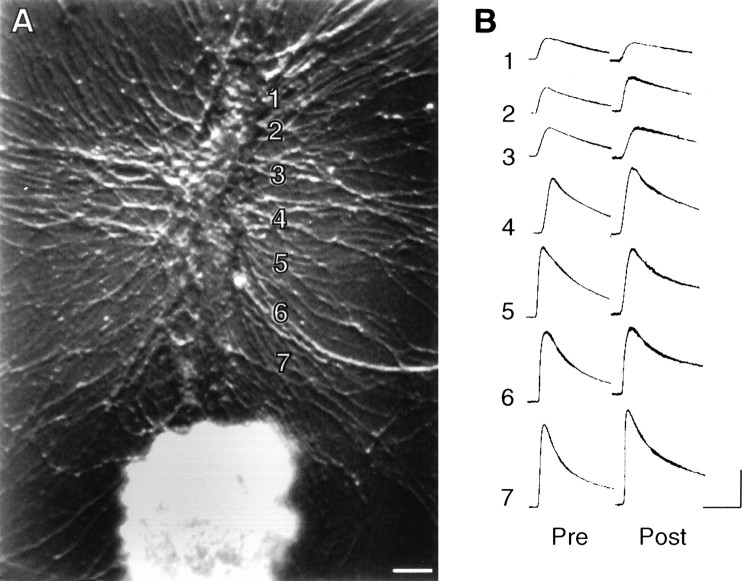
Long-term changes in the efficacy of Aplysia sensory neuron (SN) connections accompany behavioral training or applications with 5-HT. The changes evoked by training or 5-HT include formation of new SN varicosities and transmitter release sites. Because new synapse formation requires proper alignment of presynaptic structures with postsynaptic zones containing a high density of transmitter receptors, we examined whether changes in postsynaptic sensitivity to the presumed SN transmitter (glutamate) were correlated with formation and distribution of new SN varicosities in contact with motor cell L7 in cell culture. The formation of stable SN connections after 4 d in culture did not significantly change overall responses to focal applications of glutamate. However, specific sites along L7's axon apposed to SN varicosities expressed larger responses to glutamate compared with adjacent sites with few SN varicosities. After treatments with 5-HT that evoked long-term changes in both the structure and the function of SN-L7 synaptic interaction, glutamate responses increased selectively at sites along the surface of L7's axon with preexisting or new SN varicosities. Increases in postsynaptic response to glutamate 24 hr after 5-HT treatment required interaction with an SN. These results suggest that new synapse formation between neurons, either with regeneration or after external stimuli that evoke increases in synaptic efficacy, involves site-specific changes in expression of functional neurotransmitter receptors on the postsynaptic cell that is regulated by interaction with the presynaptic neuron.
Figures
References
-
- Atashi JR, Klinz SG, Ingraham CA, Matten WT, Schachner M, Maness PF. Neural cell adhesion molecules modulate tyrosine phosphorylation of tubulin in nerve growth cone membranes. Neuron. 1992;8:831–842. - PubMed
-
- Bailey CH, Chen M. Morphological basis of long-term habituation and sensitization in Aplysia. Science. 1983;220:91–93. - PubMed
-
- Bailey CH, Kander ER. Structural changes accompany memory storage. Annu Rev Physiol. 1993;55:397–426. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources