

Glutamine synthetase protects against neuronal degeneration in injured retinal tissue
- PMID: 9192685
- PMCID: PMC21278
- DOI: 10.1073/pnas.94.13.7024
Glutamine synthetase protects against neuronal degeneration in injured retinal tissue
Abstract
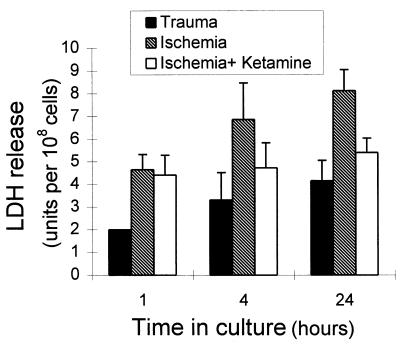
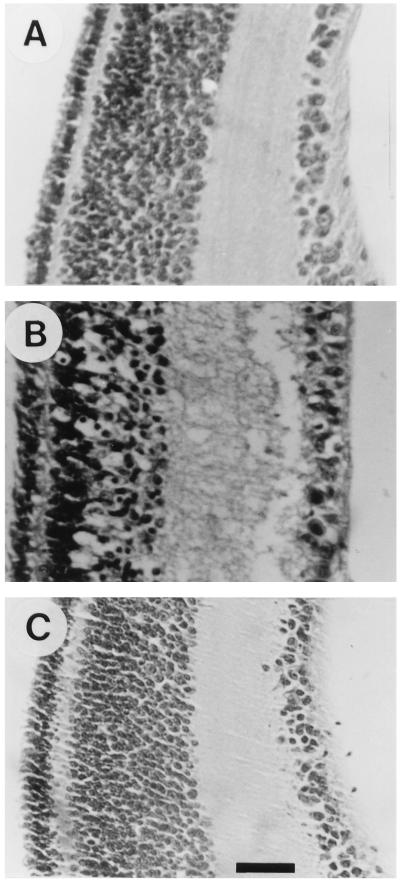
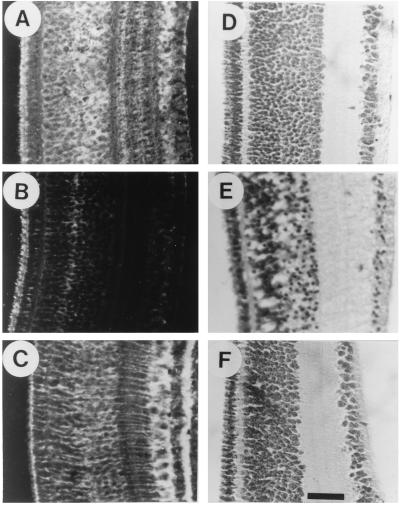
The neurotransmitter glutamate is neurotoxic when it is accumulated in a massive amount in the extracellular fluid. Excessive release of glutamate has been shown to be a major cause of neuronal degeneration after central nervous system injury. Under normal conditions, accumulation of synaptically released glutamate is prevented, at least in part, by a glial uptake system in which the glia-specific enzyme glutamine synthetase (GS) plays a key role. We postulated that glial cells cannot cope with glutamate neurotoxicity because the level of GS is not high enough to catalyze the excessive amounts of glutamate released by damaged neurons. We examined whether elevation of GS expression in glial cells protects against neuronal degeneration in injured retinal tissue. Analysis of lactate dehydrogenase efflux, DNA fragmentation, and histological sections revealed that hormonal induction of the endogenous GS gene in retinal glial cells correlates with a decline in neuronal degeneration, whereas inhibition of GS activity by methionine sulfoximine leads to increased cell death. A supply of purified GS enzyme to the culture medium of retinal explants or directly to the embryo in ovo causes a dose-dependent decline in the extent of cell death. These results show that GS is a potent neuroprotectant and that elevation of GS expression in glial cells activates an endogenous mechanism whereby neurons are protected from the deleterious effects of excess glutamate in extracellular fluid after trauma or ischemia. Our results suggest new approaches to the clinical handling of neuronal degeneration.
Figures



References
-
- Rothman S M, Olney J W. Ann Neurol. 1986;19:105–111. - PubMed
-
- Choi D W, Rothman S M. Annu Rev Neurosci. 1990;13:171–182. - PubMed
-
- Kennedy A J, Voaden M J, Marshall J. Nature (London) 1974;252:50–52. - PubMed
-
- Moscona A A. In: Progress in Retinal Research. Osborne N N, Chader G J, editors. Vol. 2. Oxford: Permagon; 1983. pp. 111–135.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
