

Chromatin dynamics in interphase nuclei and its implications for nuclear structure
- PMID: 9199163
- PMCID: PMC2137814
- DOI: 10.1083/jcb.137.7.1459
Chromatin dynamics in interphase nuclei and its implications for nuclear structure
Abstract
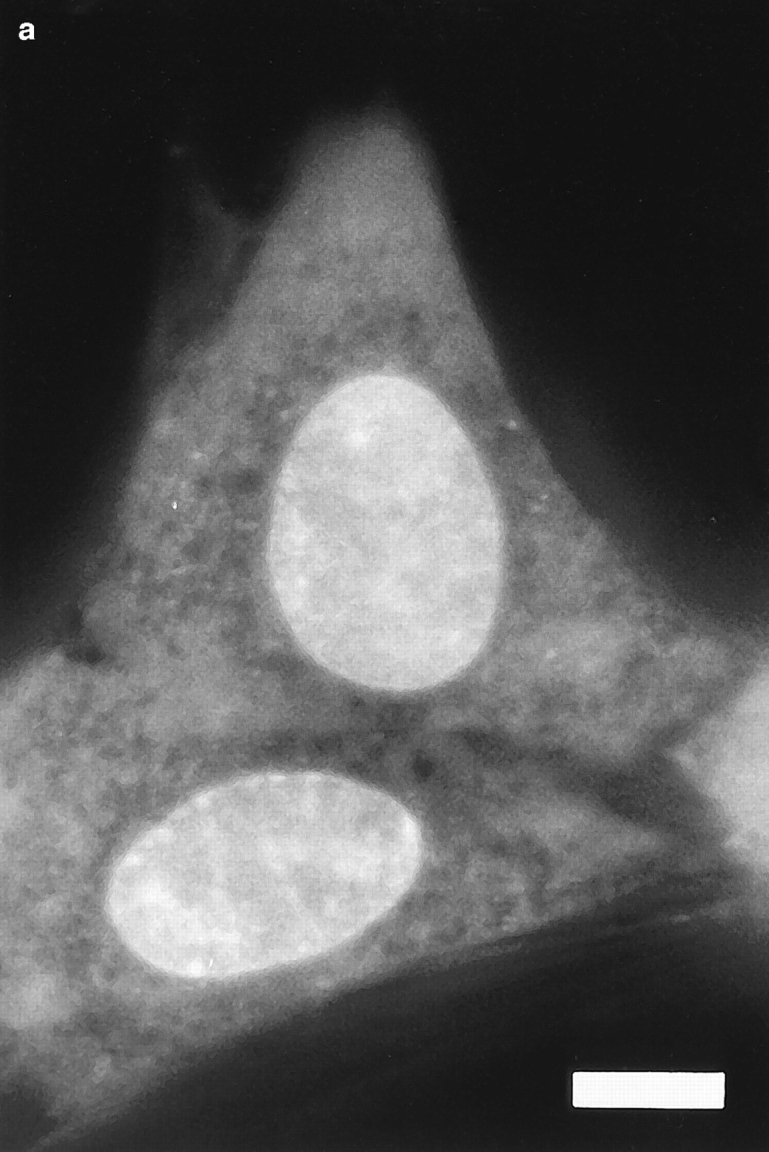
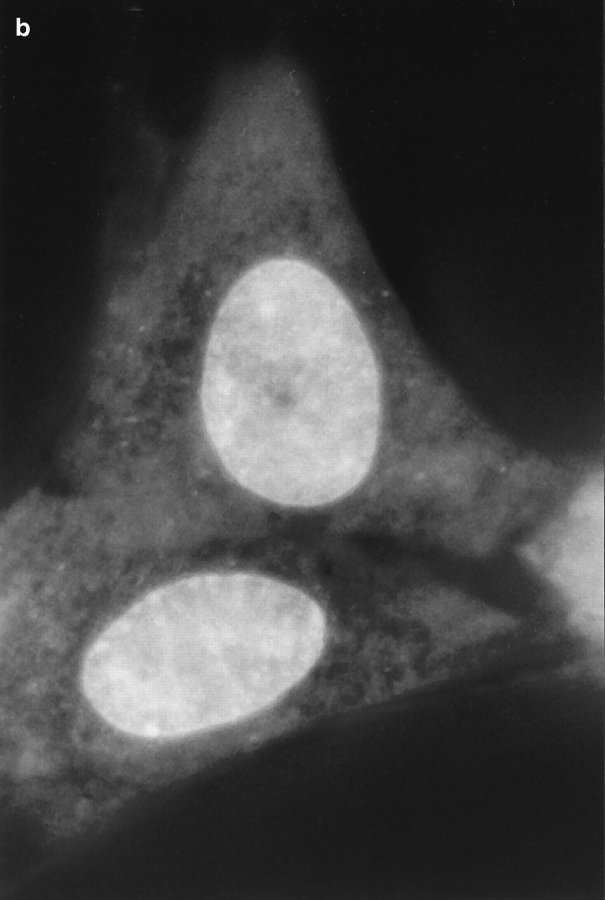
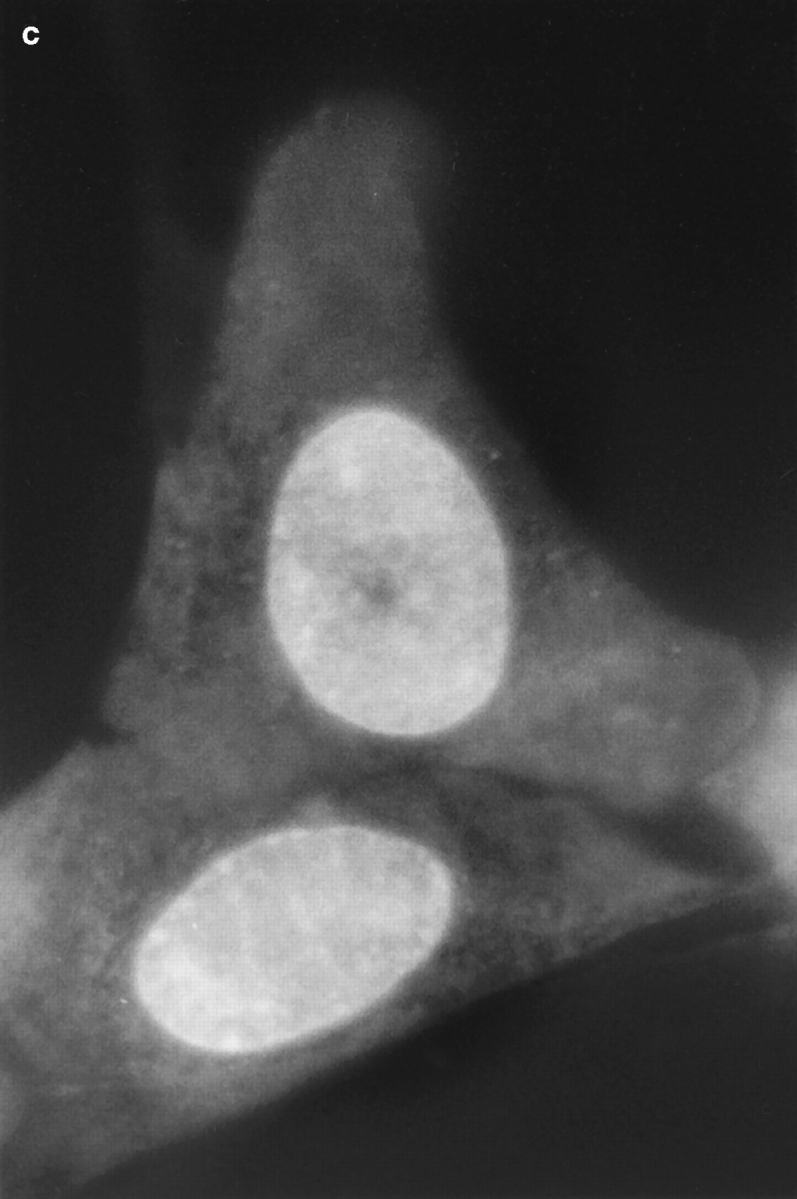
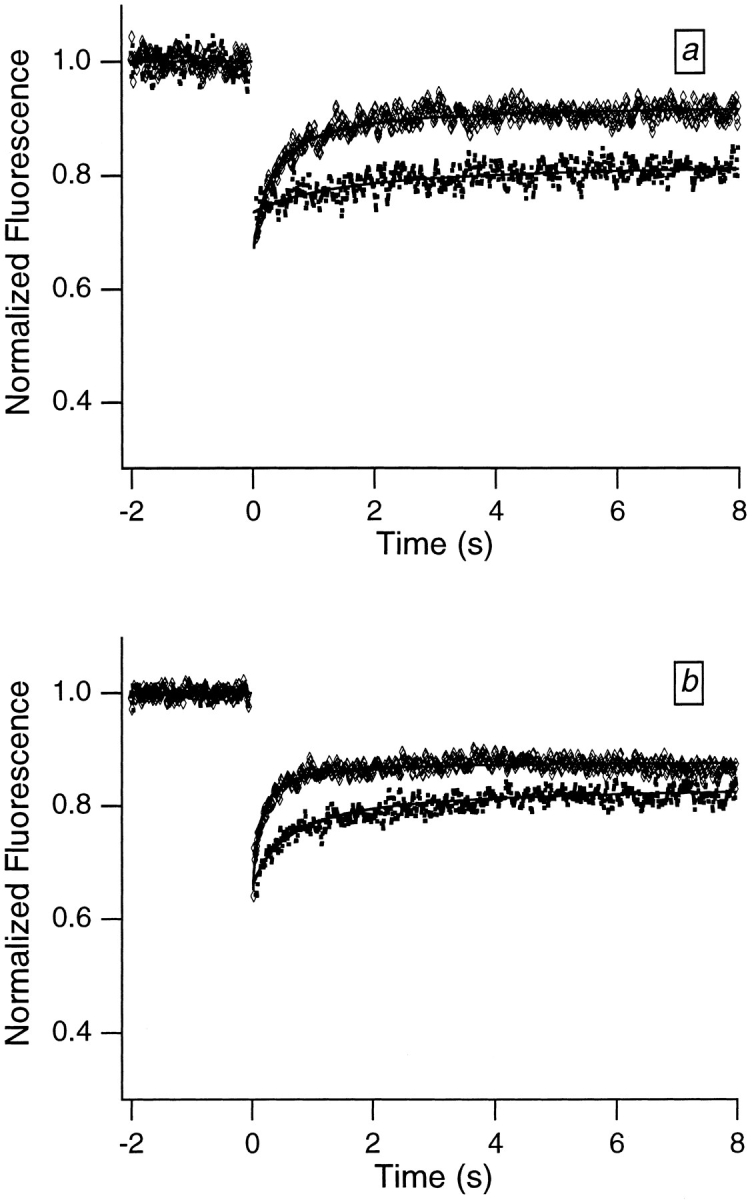
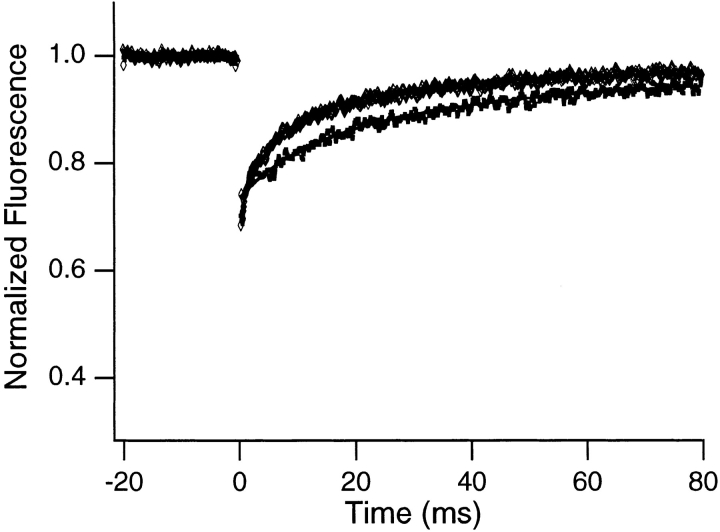
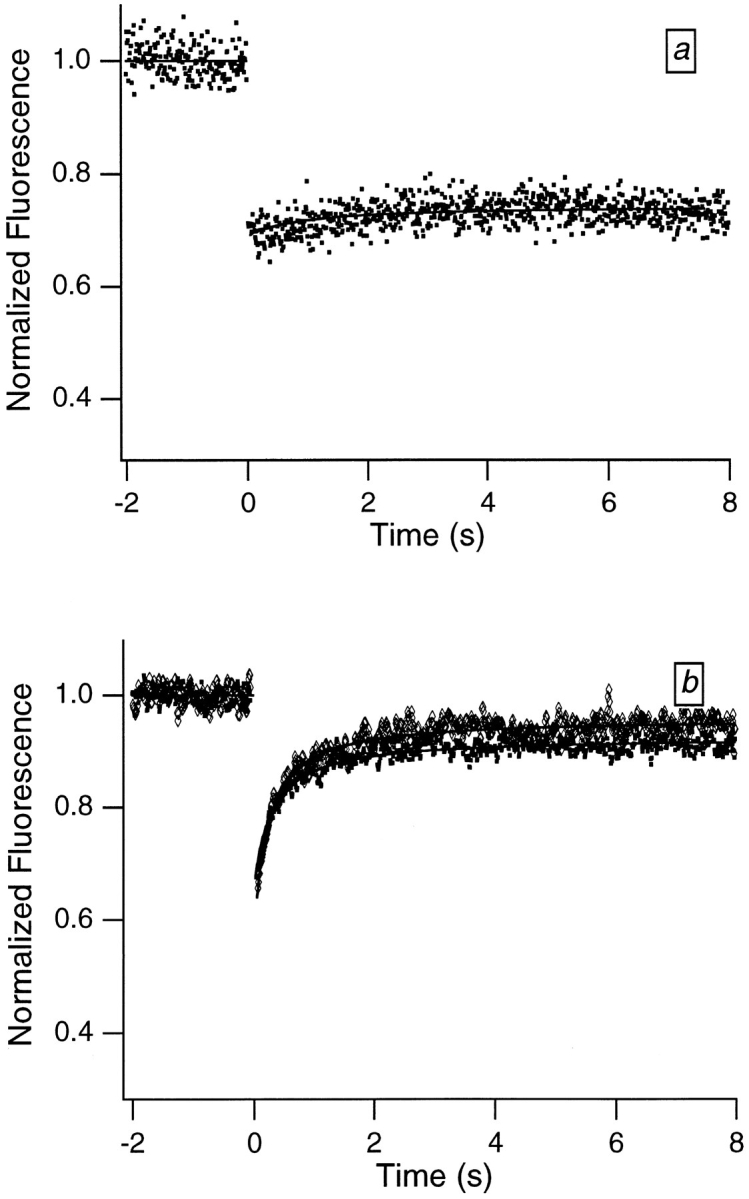
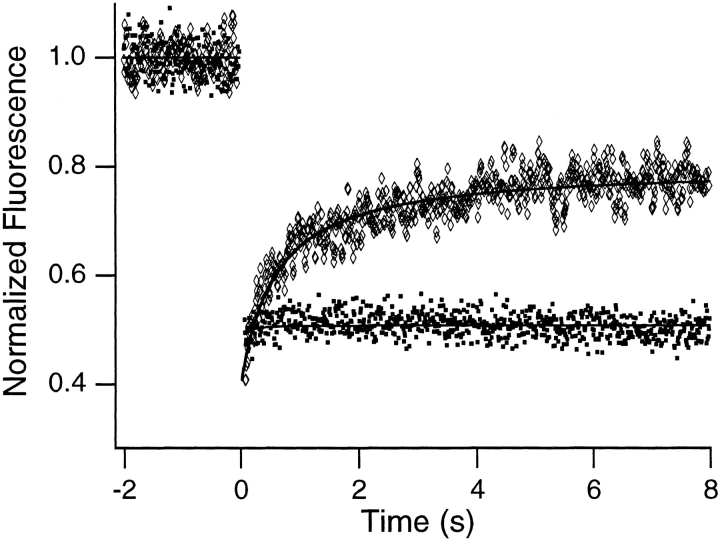
Translational dynamics of chromatin in interphase nuclei of living Swiss 3T3 and HeLa cells was studied using fluorescence microscopy and fluorescence recovery after photobleaching. Chromatin was fluorescently labeled using dihydroethidium, a membrane-permeant derivative of ethidium bromide. After labeling, a laser was used to bleach small (approximately 0.4 microm radius) spots in the heterochromatin and euchromatin of cells of both types. These spots were observed to persist for >1 h, implying that interphase chromatin is immobile over distance scales >/=0.4 microm. Over very short times (<1 s), a partial fluorescence recovery within the spots was observed. This partial recovery is attributed to independent dye motion, based on comparison with results obtained using ethidium homodimer-1, which binds essentially irreversibly to nucleic acids. The immobility observed here is consistent with chromosome confinement to domains in interphase nuclei. This immobility may reflect motion-impeding steric interactions that arise in the highly concentrated nuclear milieu or outright attachment of the chromatin to underlying nuclear substructures, such as nucleoli, the nuclear lamina, or the nuclear matrix.
Figures
References
-
- Baker, J.R. 1958. Principles of Biological Microtechnique. John Wiley & Sons Inc., New York. 357 pp.
-
- Bartholdi MF. Nuclear distribution of centromeres during the cell cycle of human diploid fibroblasts. J Cell Sci. 1991;99:255–263. - PubMed
