

The lumenal domain of Sec63p stimulates the ATPase activity of BiP and mediates BiP recruitment to the translocon in Saccharomyces cerevisiae
- PMID: 9199165
- PMCID: PMC2137819
- DOI: 10.1083/jcb.137.7.1483
The lumenal domain of Sec63p stimulates the ATPase activity of BiP and mediates BiP recruitment to the translocon in Saccharomyces cerevisiae
Abstract
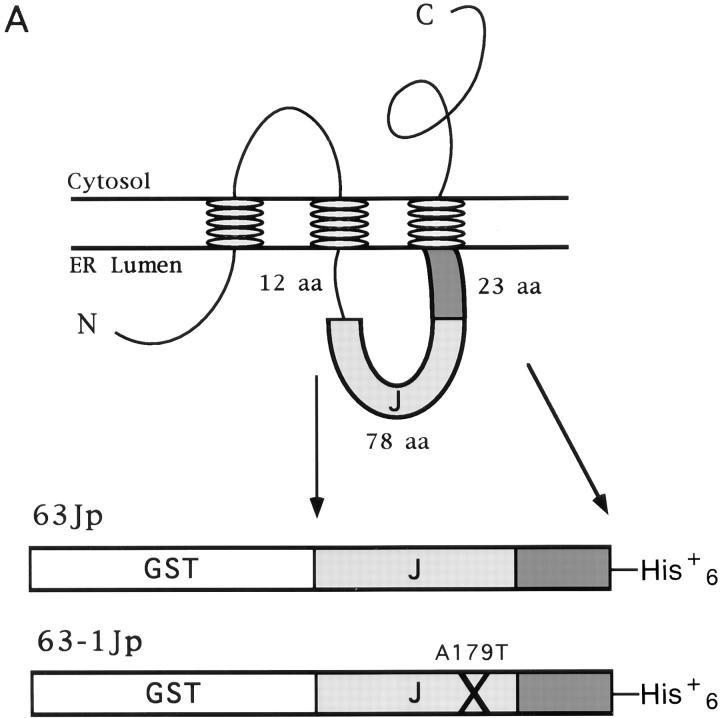
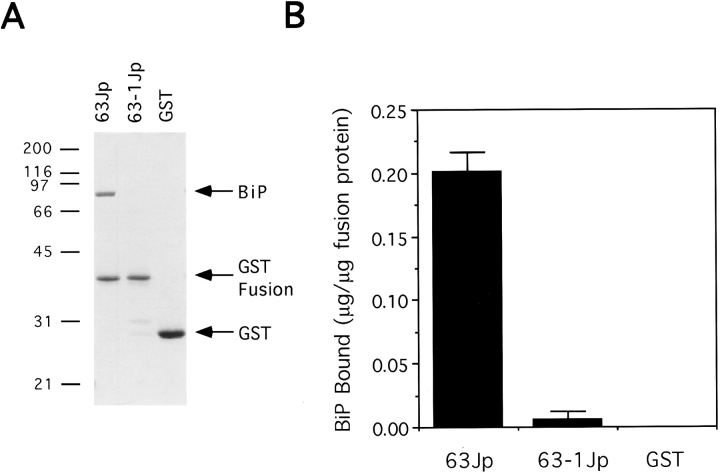
We studied the molecular nature of the interaction between the integral membrane protein Sec63p and the lumenal Hsp70 BiP to elucidate their role in the process of precursor transit into the ER of Saccharomyces cerevisiae. A lumenal stretch of Sec63p with homology to the Escherichia coli protein DnaJ is the likely region of interface between Sec63p and BiP. This domain, purified as a fusion protein (63Jp) with glutathione S-transferase (GST), mediated a stable ATP-dependent binding interaction between 63Jp and BiP and stimulated the ATPase activity of BiP. The interaction was highly selective because only BiP was retained on immobilized 63Jp when detergent-solubilized microsomes were mixed with ATP and the fusion protein. GST alone was inactive in these assays. Additionally, a GST fusion containing a point mutation in the lumenal domain of Sec63p did not interact with BiP. Finally, we found that the soluble Sec63p lumenal domain inhibited efficient precursor import into proteoliposomes reconstituted so as to incorporate both BiP and the fusion protein. We conclude that the lumenal domain of Sec63p is sufficient to mediate enzymatic interaction with BiP and that this interaction positioned at the translocation apparatus or translocon at the lumenal face of the ER is vital for protein translocation into the ER.
Figures










References
-
- Benaroudj N, Triniolles F, Ladjimi MM. Effect of nucleotides, peptides, and unfolded proteins on the self-association of the molecular chaperone HSC70. J Biol Chem. 1996;271:18471–18476. - PubMed
-
- Bernardi G. Chromatography of proteins on hydroxyapatite. Methods Enzymol. 1971;22:325–339. - PubMed
-
- Blond-Elguindi S, Fourie AM, Sambrook JF, Gething MH. Peptide-dependent stimulation of the ATPase activity of the molecular chaperone BiP is the result of conversion of oligomers to active monomers. J Biol Chem. 1993;268:12730–12735. - PubMed
-
- Brodsky JL. Post-translational protein translocation: not all hsc70s are created equal. Trends Biochem Sci. 1996;21:122–126. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
