

ATP P2X receptors mediate fast synaptic transmission in the dorsal horn of the rat spinal cord
- PMID: 9204914
- PMCID: PMC6793828
- DOI: 10.1523/JNEUROSCI.17-14-05297.1997
ATP P2X receptors mediate fast synaptic transmission in the dorsal horn of the rat spinal cord
Abstract
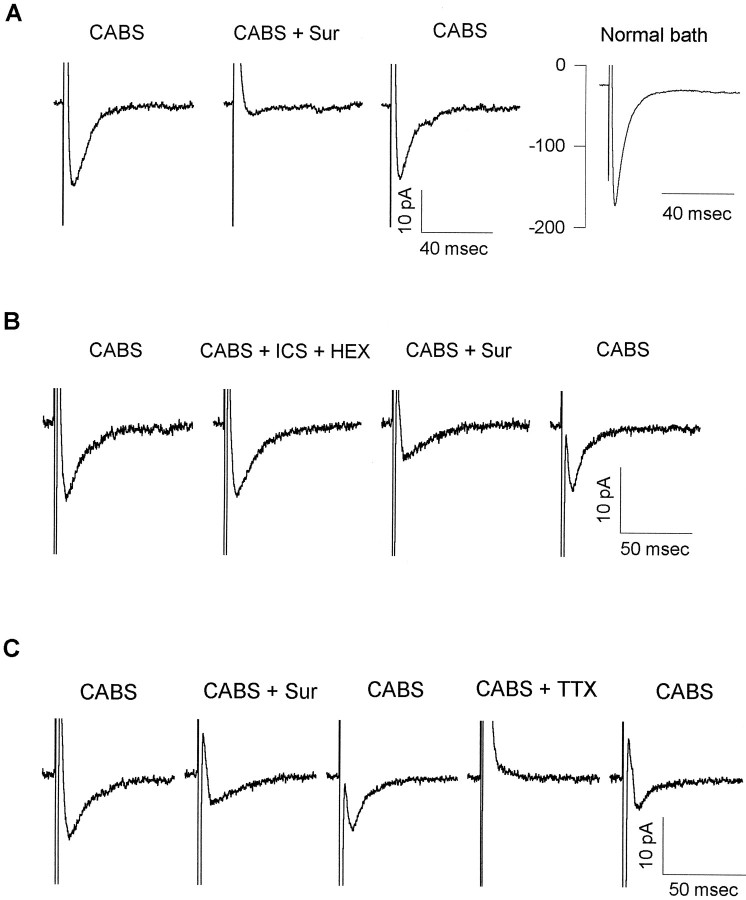
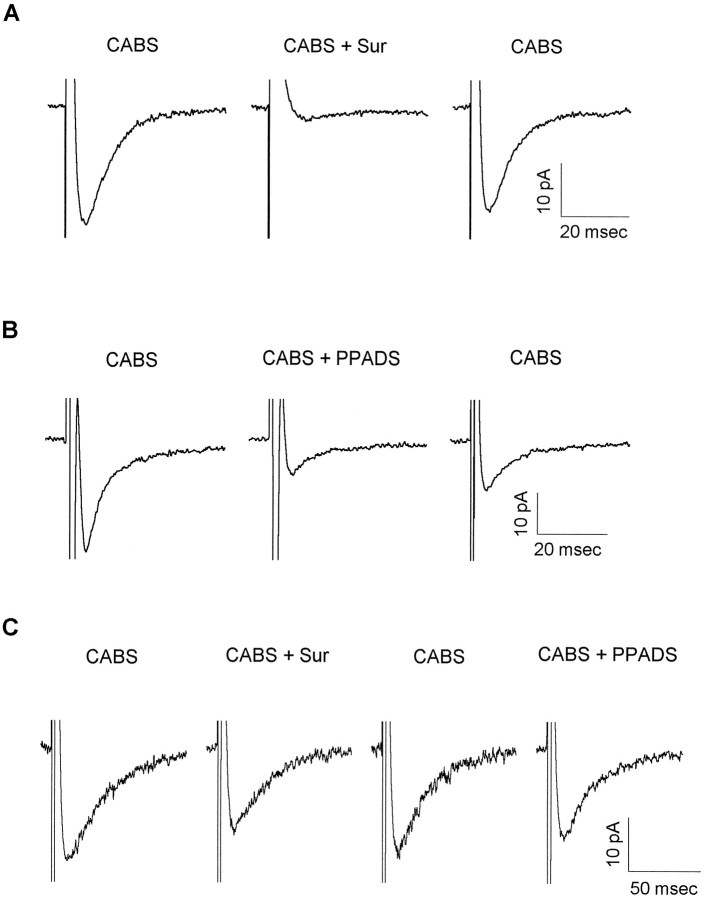
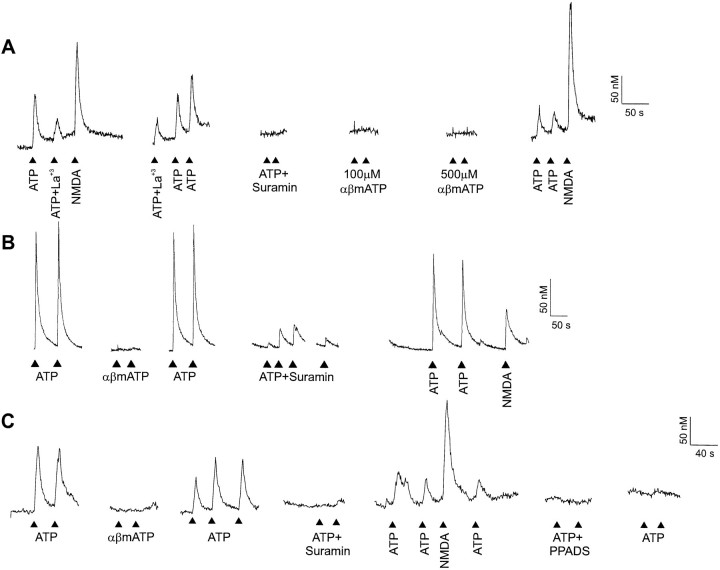
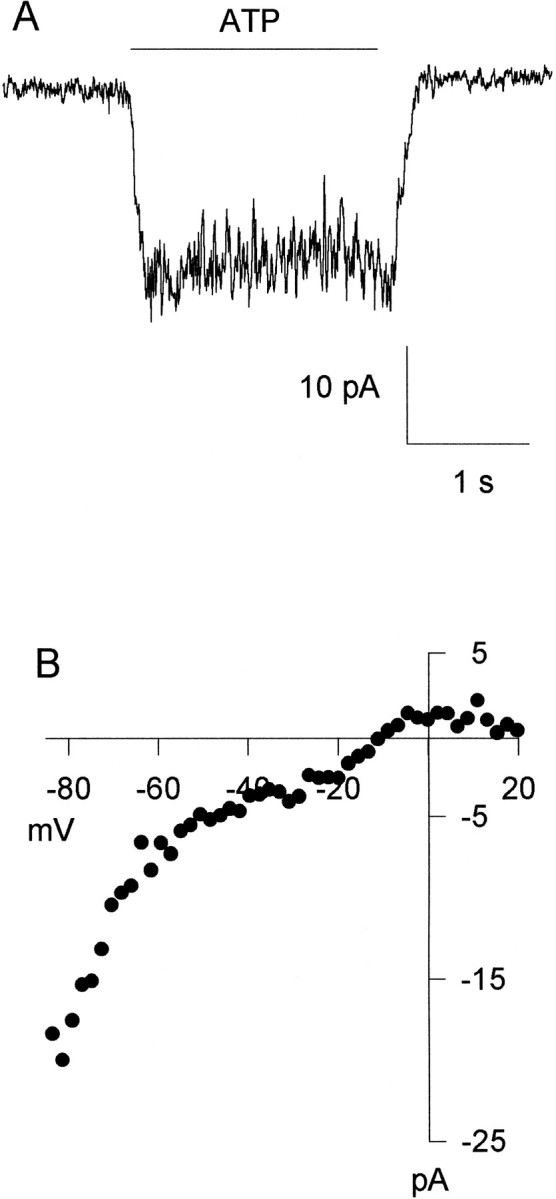
ATP has been proposed to mediate synaptic transmission in the spinal cord dorsal horn, particularly in the pathway carrying nociceptive information. Using transverse spinal cord slices from postnatal rats, we show that EPSCs mediated by P2X receptors, and presumably activated by synaptically released ATP, are evoked in a subpopulation of spinal cord lamina II neurons, a region known to receive strong input from nociceptive primary afferents. The P2X receptors on acutely dissociated dorsal horn neurons are nondesensitizing, insensitive to alphabeta methylene ATP, and show strong but variable sensitivity to the antagonists suramin and pyridoxal-phosphate-6-azophenyl-2',4'-disulfonic acid (PPADS). These characteristics are consistent with a heterogeneous population of P2X receptors, the composition of which includes P2X2, P2X4, and P2X6 receptor subtypes. Our results suggest that ATP-activated P2X receptors in lamina II of the rat spinal cord may play a role in transmitting or modulating nociceptive information.
Figures
References
-
- Bardoni R, Goldstein PA, Lee CJ, MacDermott AB. Patch-clamp recording from visually identified substantia gelatinosa neurons in postnatal rat spinal cord slices. Soc Neurosci Abstr. 1995;21:613.
-
- Benham CD, Tsien RW. A novel receptor-operated Ca2+-permeable channel activated by ATP in smooth muscle. Nature. 1987;328:275–278. - PubMed
-
- Brake AJ, Wagenbach MJ, Julius D. New structural motif for ligand-gated ion channels defined by an ionotropic ATP receptor. Nature. 1994;371:519–523. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources