

Identification of the yeast 20S proteasome catalytic centers and subunit interactions required for active-site formation
- PMID: 9207060
- PMCID: PMC23776
- DOI: 10.1073/pnas.94.14.7156
Identification of the yeast 20S proteasome catalytic centers and subunit interactions required for active-site formation
Abstract
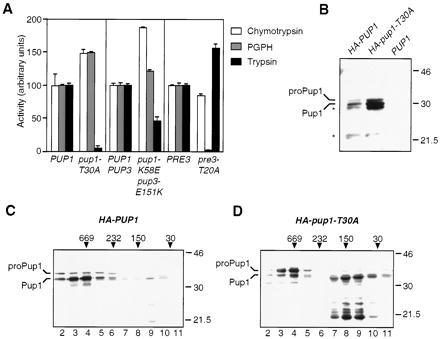
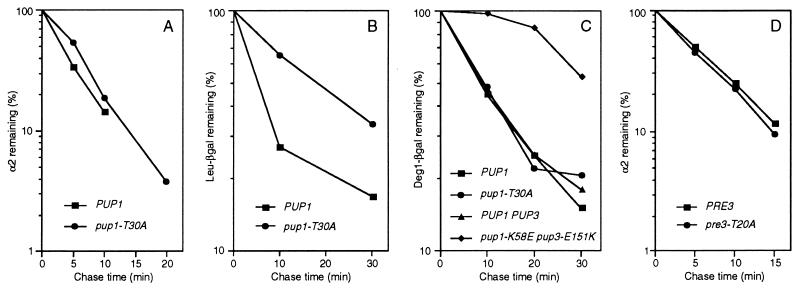
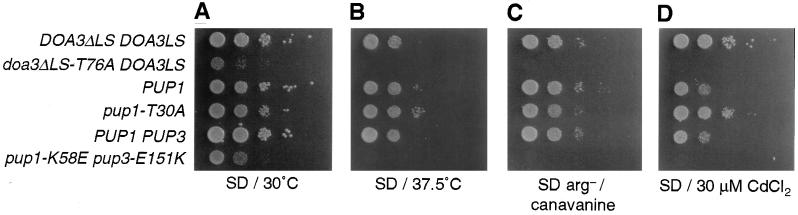
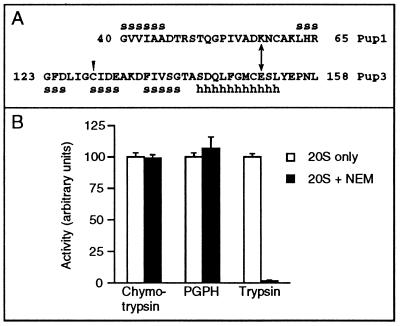
The proteasome is responsible for degradation of substrates of the ubiquitin pathway. 20S proteasomes are cylindrical particles with subunits arranged in a stack of four heptameric rings. The outer rings are composed of alpha subunits, and the inner rings are composed of beta subunits. A well-characterized archaeal proteasome has a single type of each subunit, and the N-terminal threonine of the beta subunit is the active-site nucleophile. Yeast proteasomes have seven different beta subunits and exhibit several distinct peptidase activities, which were proposed to derive from disparate active sites. We show that mutating the N-terminal threonine in the yeast Pup1 beta subunit eliminates cleavage after basic residues in peptide substrates, and mutating the corresponding threonine of Pre3 prevents cleavage after acidic residues. Surprisingly, neither mutation has a strong effect on cell growth, and they have at most minor effects on ubiquitin-dependent proteolysis. We show that Pup1 interacts with Pup3 in each beta subunit ring. Our data reveal that different proteasome active sites contribute very differently to protein breakdown in vivo, that contacts between particular subunits in each beta subunit ring are critical for active-site formation, and that active sites in archaea and different eukaryotes are highly similar.
Figures

References
-
- Hochstrasser M. Annu Rev Genet. 1996;30:405–439. - PubMed
-
- Rubin D M, Finley D. Curr Biol. 1995;5:854–858. - PubMed
-
- Löwe J, Stock D, Jap B, Zwickl P, Baumeister W, Huber R. Science. 1995;268:533–539. - PubMed
-
- Seemüller E, Lupas A, Stock D, Löwe J, Huber R, Baumeister W. Science. 1995;268:579–582. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
