

Interaction between replication protein A and p53 is disrupted after UV damage in a DNA repair-dependent manner
- PMID: 9207066
- PMCID: PMC23787
- DOI: 10.1073/pnas.94.14.7186
Interaction between replication protein A and p53 is disrupted after UV damage in a DNA repair-dependent manner
Abstract
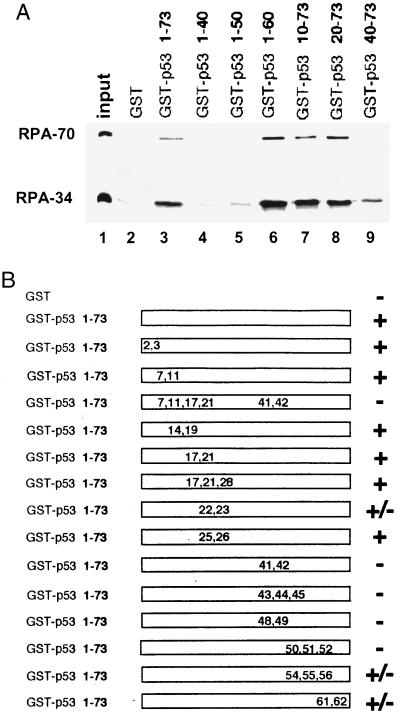
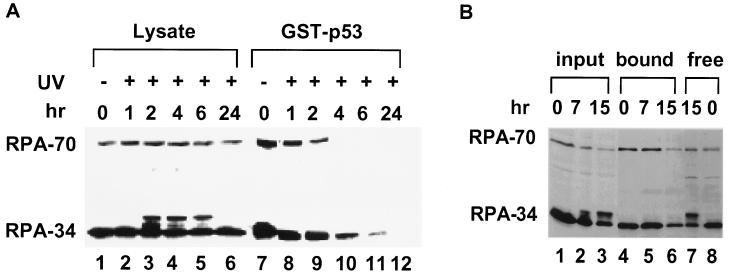
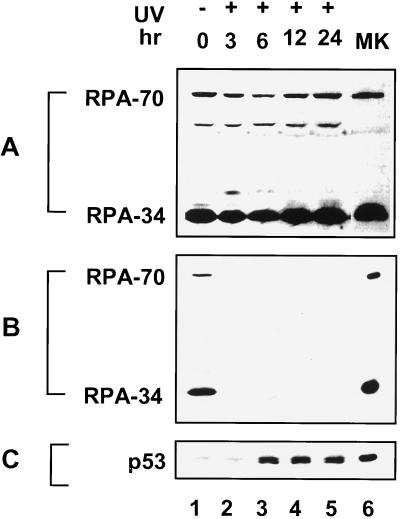
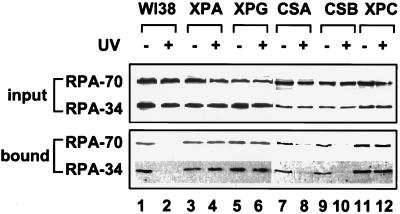
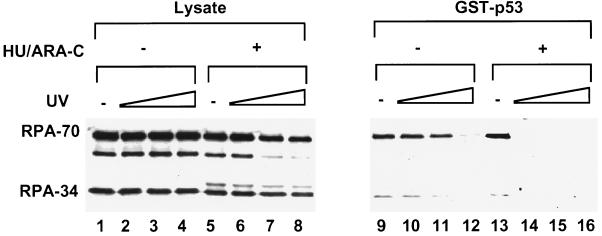
Replication protein A (RPA) is required for both DNA replication and nucleotide excision repair. Previous studies have shown that RPA interacts with the tumor suppressor p53. Herein, we have mapped a 20-amino acid region in the N-terminal part of p53 that is essential for its binding to RPA. This region is distinct from the minimal activation domain of p53 previously identified. We also demonstrate that UV radiation of cells greatly reduces the ability of RPA to bind to p53. Interestingly, damage-induced hyperphosphorylated RPA does not associate with p53. Furthermore, down-regulation of the RPA/p53 interaction is dependent upon the capability of cells to perform global genome repair. On the basis of these data, we propose that RPA may participate in the coordination of DNA repair with the p53-dependent checkpoint control by sensing UV damage and releasing p53 to activate its downstream targets.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
