

Distinct roles for signals relayed through the common cytokine receptor gamma chain and interleukin 7 receptor alpha chain in natural T cell development
- PMID: 9221763
- PMCID: PMC2198975
- DOI: 10.1084/jem.186.2.331
Distinct roles for signals relayed through the common cytokine receptor gamma chain and interleukin 7 receptor alpha chain in natural T cell development
Abstract
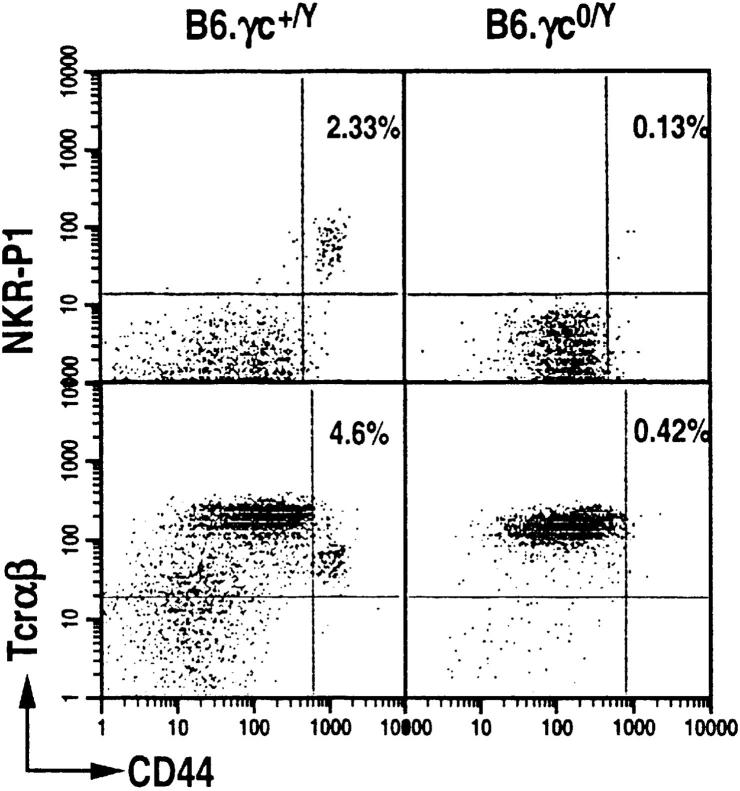
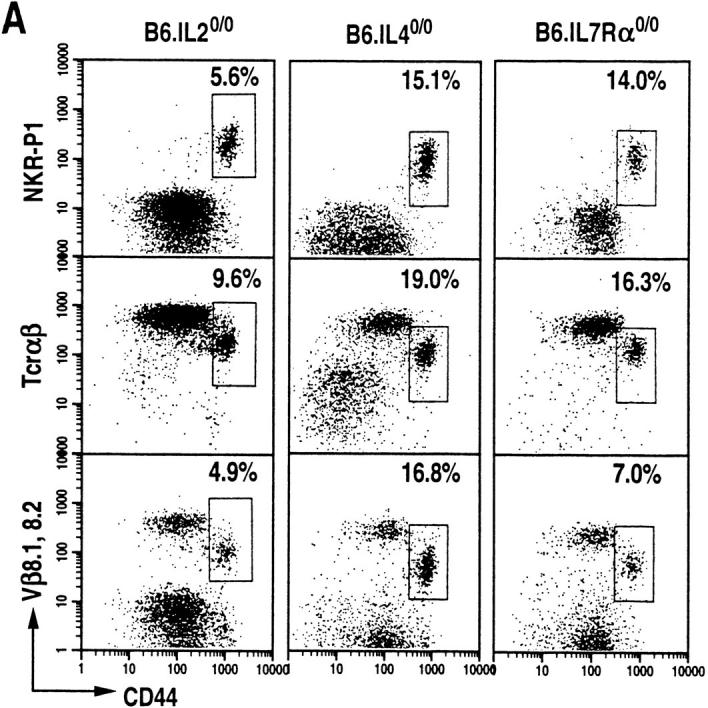
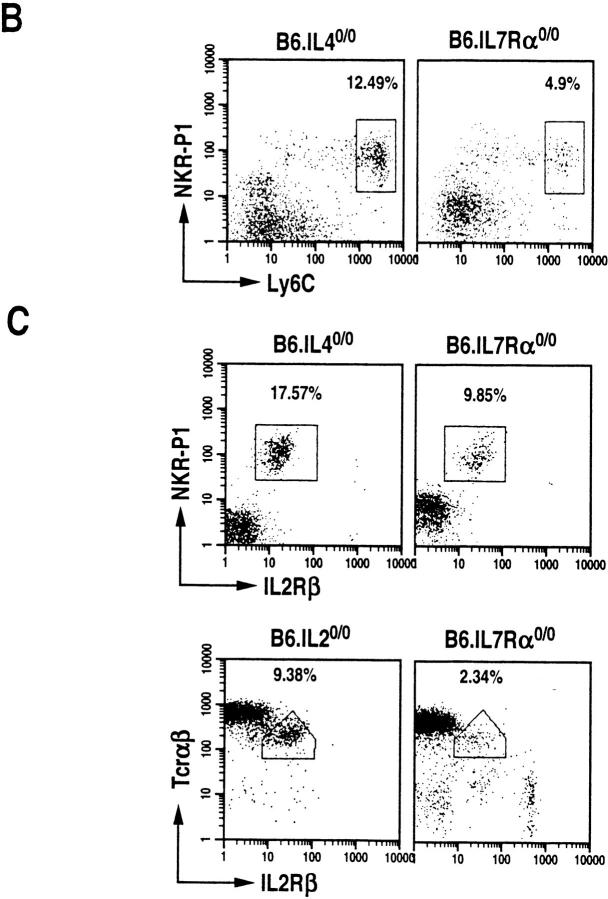
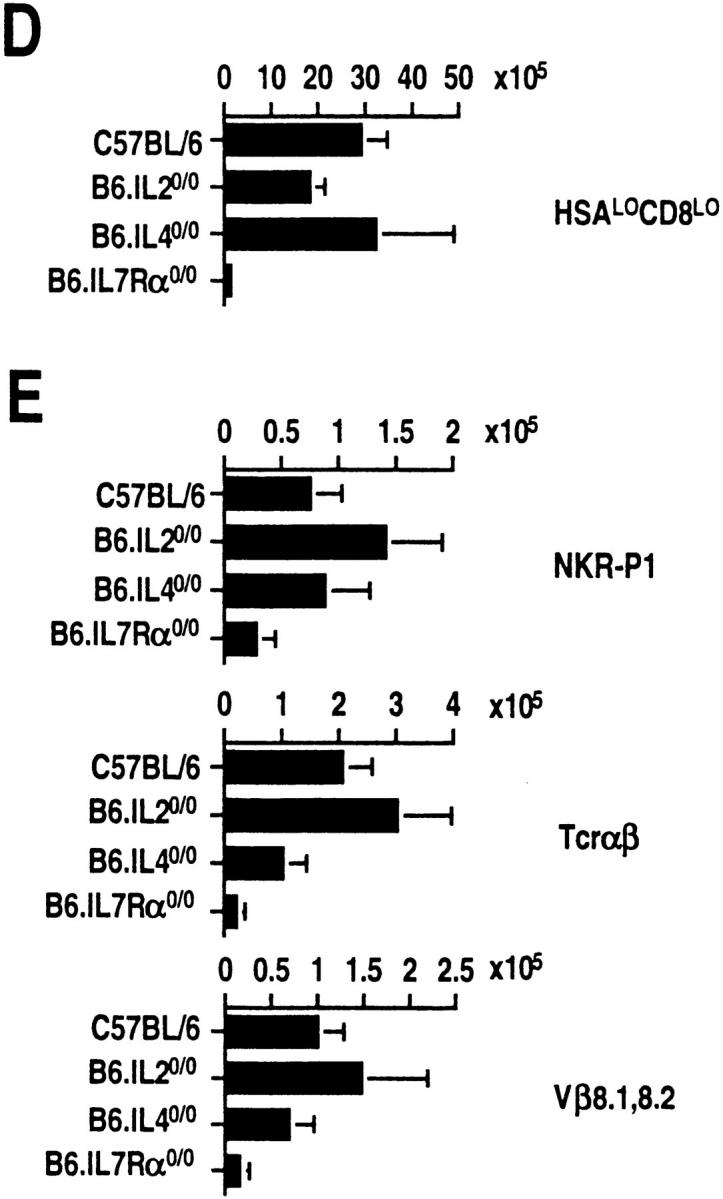
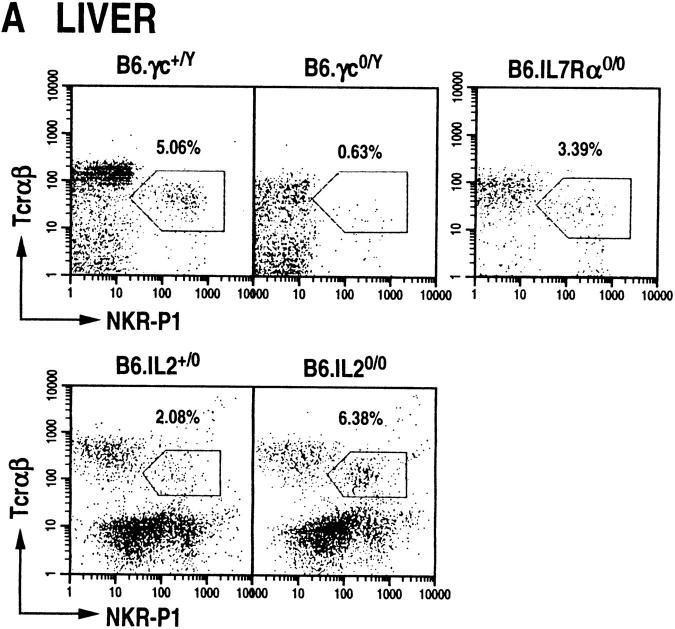
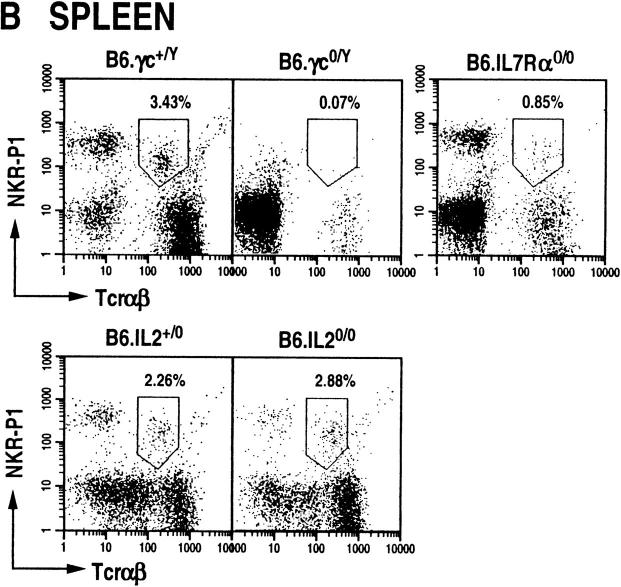
The commitment, differentiation, and expansion of mainstream alpha/beta T cells during ontogeny depend on the highly controlled interplay of signals relayed by cytokines through their receptors on progenitor cells. The role of cytokines in the development of natural killer (NK)1(+) natural T cells is less clearly understood. In an approach to define the role of cytokines in the commitment, differentiation, and expansion of NK1(+) T cells, their development was studied in common cytokine receptor gamma chain (gammac) and interleukin (IL)-7 receptor alpha (IL-7Ralpha)-deficient mice. These mutations block mainstream alpha/beta T cell ontogeny at an early prethymocyte stage. Natural T cells do not develop in gammac-deficient mice; they are absent in the thymus and peripheral lymphoid organs such as the liver and the spleen. In contrast, NK1(+) T cells develop in IL-7Ralpha-deficient mice in the thymus, and they are present in the liver and in the spleen. However, the absolute number of NK1(+) T cells in the thymus of IL-7Ralpha-deficient mice is reduced to approximately 10%, compared to natural T cell number in the wild-type thymus. Additional data revealed that NK1(+) T cell ontogeny is not impaired in IL-2- or IL-4-deficient mice, suggesting that neither IL-2, IL-4, nor IL-7 are required for their development. From these data, we conclude that commitment and/or differentiation to the NK1(+) natural T cell lineage requires signal transduction through the gammac, and once committed, their expansion requires signals relayed through the IL-7Ralpha.
Figures
References
-
- Vicari AP, Zlotnik A. Mouse NK1.1+T cells: a new family of T cells. Immunol Today. 1996;17:71–76. - PubMed
-
- Smiley ST, Kaplan MH, Grusby MJ. Immunoglobulin E production in the absence of interleukin-4 secreting CD1 dependent cells. Science (Wash DC) 1997;275:977–979. - PubMed
-
- Chen Y-H, Chiu NM, Mandal M, Wang N, Wang C-R. Impaired NK1+T cell development and early IL-4 production in CD1 deficient mice. Immunity. 1997;6:459–467. - PubMed
-
- Mendiratta SK, Martin WD, Hong S, Boesteanu A, Joyce S, Van Kaer L. CD1d1mutant mice are deficient in natural T cells that promptly produce IL-4. Immunity. 1997;6:469–477. - PubMed
-
- Joyce S, Negishi I, Boesteanu A, DeSilva AD, Sharma P, Chorney MJ, Loh DY, Van Kaer L. Expansion of natural (NK1+) T cells that express αβ T cell receptors in transporters associated with antigen presentation-1 null and thymus leukemia antigen positive mice. J Exp Med. 1996;184:1579–1584. - PMC - PubMed
