

An important developmental role for oligosaccharides during early embryogenesis of cyprinid fish
- PMID: 9223299
- PMCID: PMC21541
- DOI: 10.1073/pnas.94.15.7982
An important developmental role for oligosaccharides during early embryogenesis of cyprinid fish
Abstract
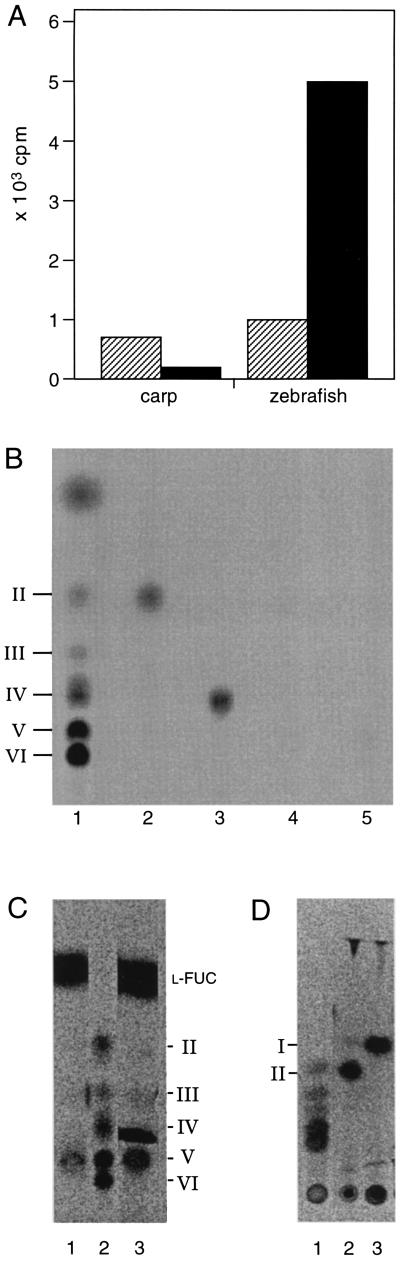
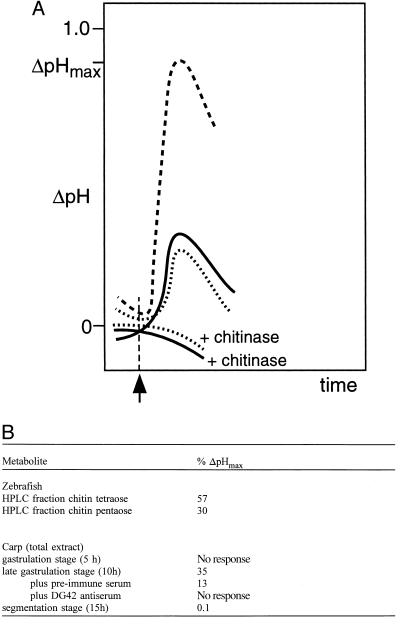
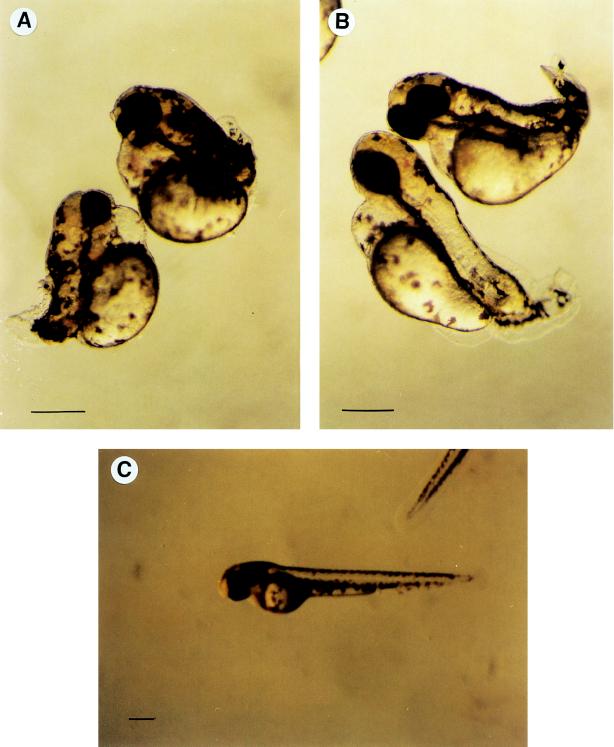
Derivatives of chitin oligosaccharides have been shown to play a role in plant organogenesis at nanomolar concentrations. Here we present data which indicate that chitin oligosaccharides are important for embryogenesis in vertebrates. We characterize chitin oligosaccharides synthesized in vitro by zebrafish and carp embryos in the late gastrulation stage by incorporation of radiolabeled N-acetyl-D-[U14C]glucosamine and by HPLC in combination with enzymatic conversion using the Bradyrhizobium NodZ alpha-1, 6-fucosyltransferase and chitinases. A rapid and sensitive bioassay for chitin oligosaccharides was also used employing suspension-cultured plant cells of Catharanthus roseus. We show that chitin oligosaccharide synthase activity is apparent only during late gastrulation and can be inhibited by antiserum raised against the Xenopus DG42 protein. The DG42 protein, a glycosyltransferase, is transiently expressed between midblastula and neurulation in Xenopus and zebrafish embryogenesis. Microinjection of the DG42 antiserum or the Bradyrhizobium NodZ enzyme in fertilized eggs of zebrafish led to severe defects in trunk and tail development.
Figures
References
-
- Lerouge P, Roche P, Faucher C, Maillet F, Truchet G, Promé J C, Dénarié J. Nature (London) 1990;344:781–784. - PubMed
-
- Spaink H P, Sheeley D M, van Brussel A A N, Glushka J, York W S, Tak T, Geiger O, Kennedy E P, Reinhold V N, Lugtenberg B J J. Nature (London) 1991;354:125–130. - PubMed
-
- van Brussel A A N, Bakhuizen R, van Spronsen P C, Spaink H P, Tak T, Lugtenberg B J J, Kijne J W. Science. 1992;257:70–72. - PubMed
-
- Spaink H P. Annu Rev Phytopathol. 1995;33:345–368. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
