

Transitions in the coupling of transcription and nucleotide excision repair within RNA polymerase II-transcribed genes of Saccharomyces cerevisiae
- PMID: 9223308
- PMCID: PMC21550
- DOI: 10.1073/pnas.94.15.8027
Transitions in the coupling of transcription and nucleotide excision repair within RNA polymerase II-transcribed genes of Saccharomyces cerevisiae
Abstract
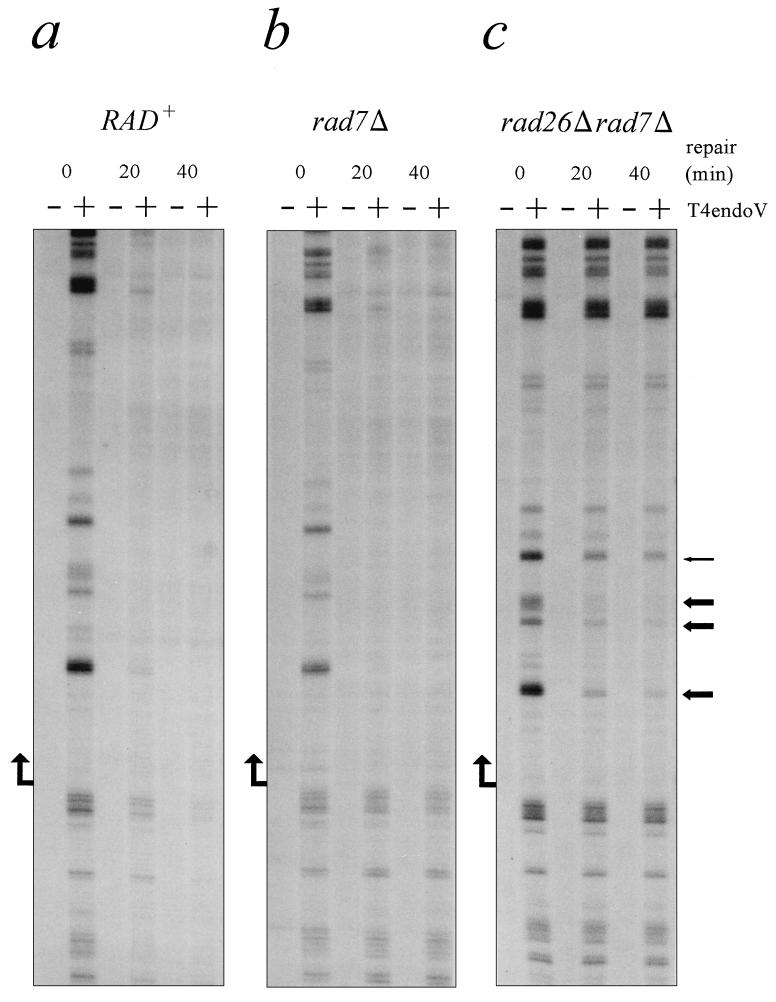
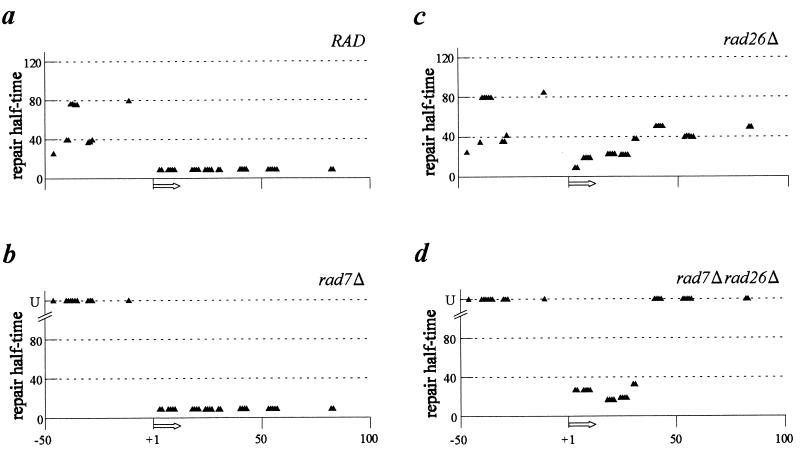
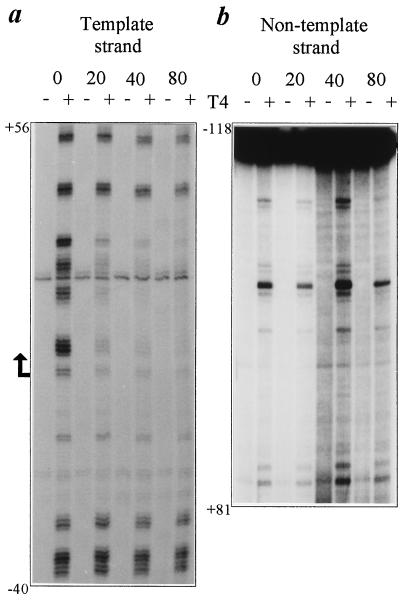
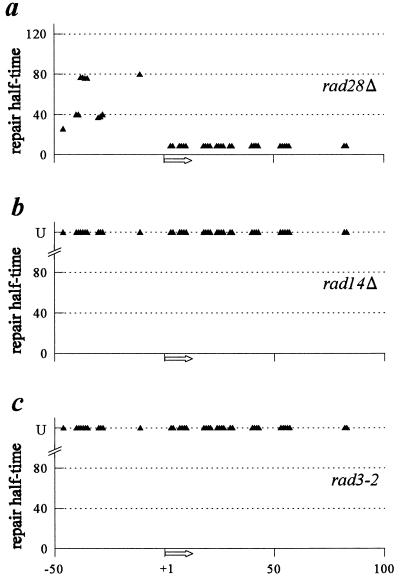
The molecular mechanism of transcription-coupled nucleotide excision repair in eukaryotes is poorly understood. The identification of the dual role of basal transcription factor TFIIH in DNA repair and transcription provided a plausible link between both processes. However, TFIIH is not part of the elongating transcription complex, suggesting that additional components are required to recruit TFIIH when RNA polymerase II (RNAPII) stalls at the site of DNA damage. Previously, we have shown that the yeast Rad26 protein is involved in transcription-coupled DNA repair. This paper describes the differential contribution of the Rad26 protein to efficient removal of UV-induced cyclobutane pyrimidine dimers (CPDs) from transcribed DNA. Two distinct regions within the transcribed strand of RNAPII-transcribed genes are identified that differ in their requirement for the RAD26 gene product. Using high-resolution repair analysis, we determined the in vivo repair kinetics of cyclobutane pyrimidine dimers positioned around the transcription initiation site of RNAPII-transcribed genes RPB2 and URA3. Although transcription-coupled repair is severely reduced in rad26 mutants, lesions positioned in a small region immediately downstream of transcription initiation are efficiently removed in the absence of Rad26. The observed transition in repair characteristics is abrupt and in excellent agreement with the region where TFIIH dissociates from RNAPII in vitro, strongly suggesting an inverse correlation between TFIIH association and Rad26 requirement. These data suggest that a transcription repair coupling factor (Rad26/CSB) is required for efficient repair only during the elongating stages of RNAPII transcription.
Figures
References
-
- Mellon I, Hanawalt P C. Nature (London) 1989;342:95–98. - PubMed
-
- Smerdon M J, Thoma F. Cell. 1990;61:675–684. - PubMed
-
- Mellon I M, Spivak G S, Hanawalt P C. Cell. 1987;51:241–249. - PubMed
-
- Selby C P, Sancar A. J Biol Chem. 1990;265:21330–21336. - PubMed
-
- Selby C P, Sancar A. Science. 1993;260:53–58. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
