

The hotspot conversion paradox and the evolution of meiotic recombination
- PMID: 9223314
- PMCID: PMC21556
- DOI: 10.1073/pnas.94.15.8058
The hotspot conversion paradox and the evolution of meiotic recombination
Abstract
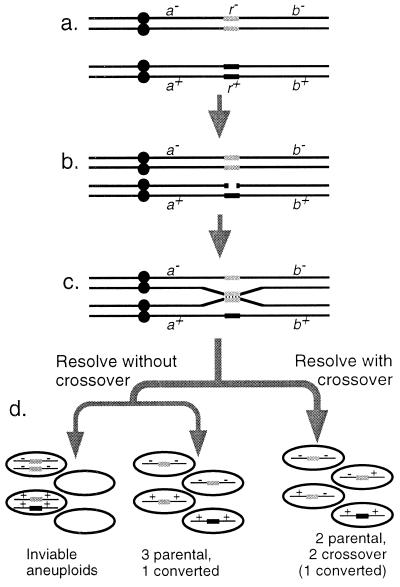
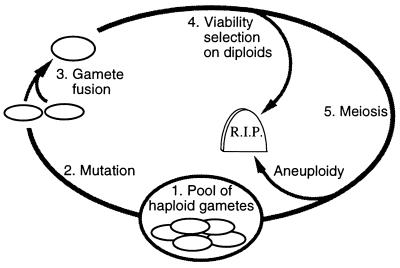
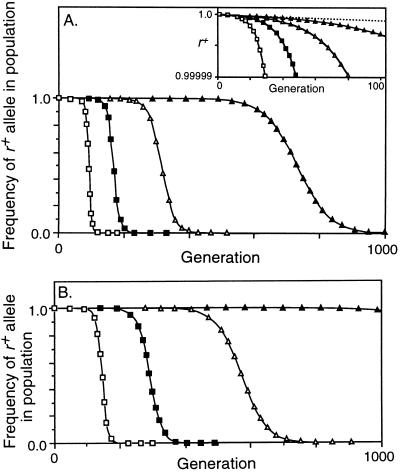
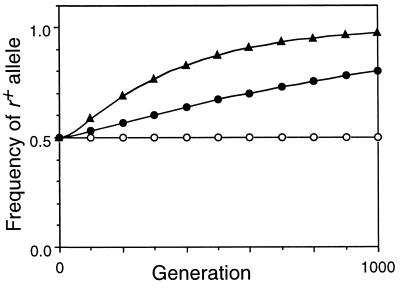
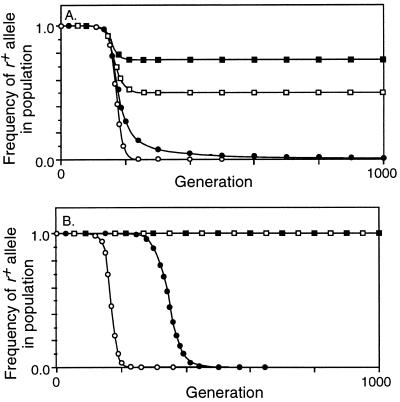
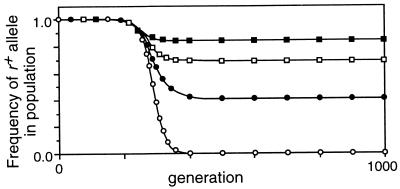
Studies of meiotic recombination have revealed an evolutionary paradox. Molecular and genetic analysis has shown that crossing over initiates at specific sites called hotspots, by a recombinational-repair mechanism in which the initiating hotspot is replaced by a copy of its homolog. We have used computer simulations of large populations to show that this mechanism causes active hotspot alleles to be rapidly replaced by inactive alleles, which arise by rare mutation and increase by recombination-associated conversion. Additional simulations solidified the paradox by showing that the known benefits of recombination appear inadequate to maintain its mechanism. Neither the benefits of accurate segregation nor those of recombining flanking genes were sufficient to preserve active alleles in the face of conversion. A partial resolution to this paradox was obtained by introducing into the model an additional, nonmeiotic function for the sites that initiate recombination, consistent with the observed association of hotspots with functional sites in chromatin. Provided selection for this function was sufficiently strong, active hotspots were able to persist in spite of frequent conversion to inactive alleles. However, this explanation is unsatisfactory for two reasons. First, it is unlikely to apply to obligately sexual species, because observed crossover frequencies imply maintenance of many hotspots per genome, and the viability selection needed to preserve these would drive the species to extinction. Second, it fails to explain why such a genetically costly mechanism of recombination has been maintained over evolutionary time. Thus the paradox persists and is likely to be resolved only by significant changes to the commonly accepted mechanism of crossing over.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
