

Mouse model of GM2 activator deficiency manifests cerebellar pathology and motor impairment
- PMID: 9223328
- PMCID: PMC21570
- DOI: 10.1073/pnas.94.15.8138
Mouse model of GM2 activator deficiency manifests cerebellar pathology and motor impairment
Abstract
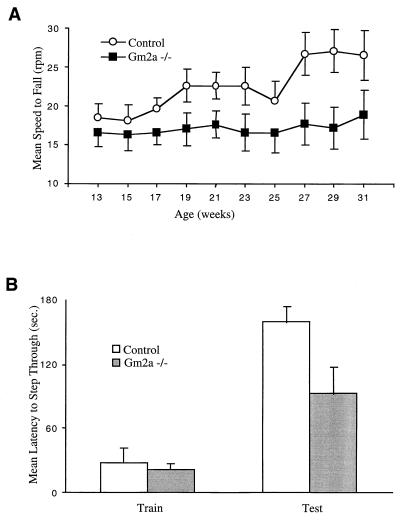
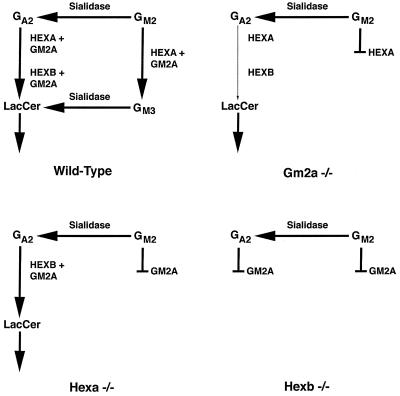
The GM2 activator deficiency (also known as the AB variant), Tay-Sachs disease, and Sandhoff disease are the major forms of the GM2 gangliosidoses, disorders caused by defective degradation of GM2 ganglioside. Tay-Sachs and Sandhoff diseases are caused by mutations in the genes (HEXA and HEXB) encoding the subunits of beta-hexosaminidase A. The GM2 activator deficiency is caused by mutations in the GM2A gene encoding the GM2 activator protein. For degradation of GM2 ganglioside by beta-hexosamindase A, the GM2 activator protein must participate by forming a soluble complex with the ganglioside. In each of the disorders, GM2 ganglioside and related lipids accumulate to pathologic levels in neuronal lysosomes, resulting in clinically similar disorders with an onset in the first year of life, progressive neurodegeneration, and death by early childhood. We previously have described mouse models of Tay-Sachs (Hexa -/-) and Sandhoff (Hexb -/-) diseases with vastly different clinical phenotypes. The Hexa -/- mice were asymptomatic whereas the Hexb -/- mice were severely affected. Through gene disruption in embryonic stem cells we now have established a mouse model of the GM2 activator deficiency that manifests an intermediate phenotype. The Gm2a -/- mice demonstrated neuronal storage but only in restricted regions of the brain (piriform, entorhinal cortex, amygdala, and hypothalamic nuclei) reminiscent of the asymptomatic Tay-Sachs model mice. However, unlike the Tay-Sachs mice, the Gm2a -/- mice displayed significant storage in the cerebellum and defects in balance and coordination. The abnormal ganglioside storage in the Gm2a -/- mice consisted of GM2 with a low amount of GA2. The results demonstrate that the activator protein is required for GM2 degradation and also may indicate a role for the GM2 activator in GA2 degradation.
Figures
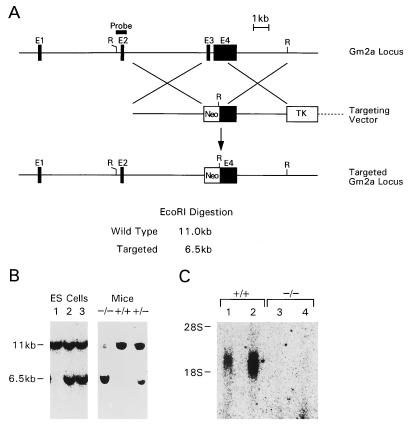
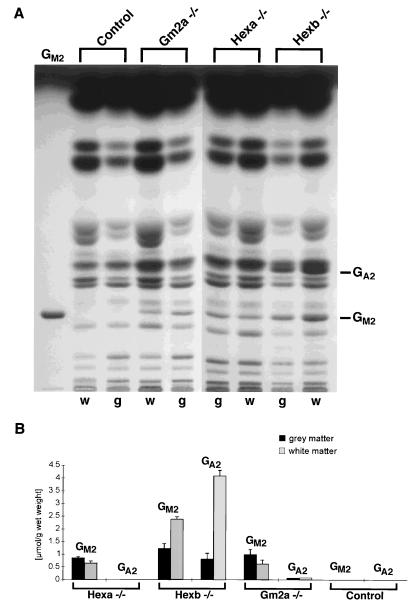
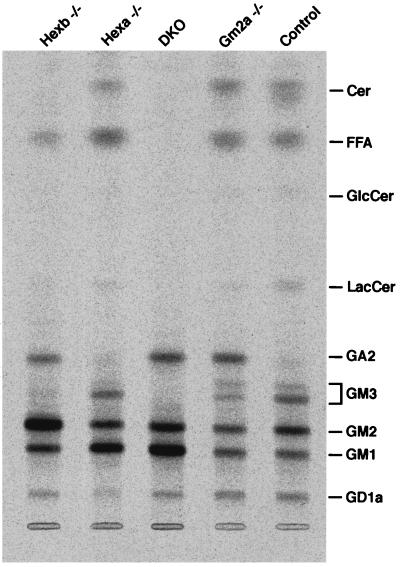
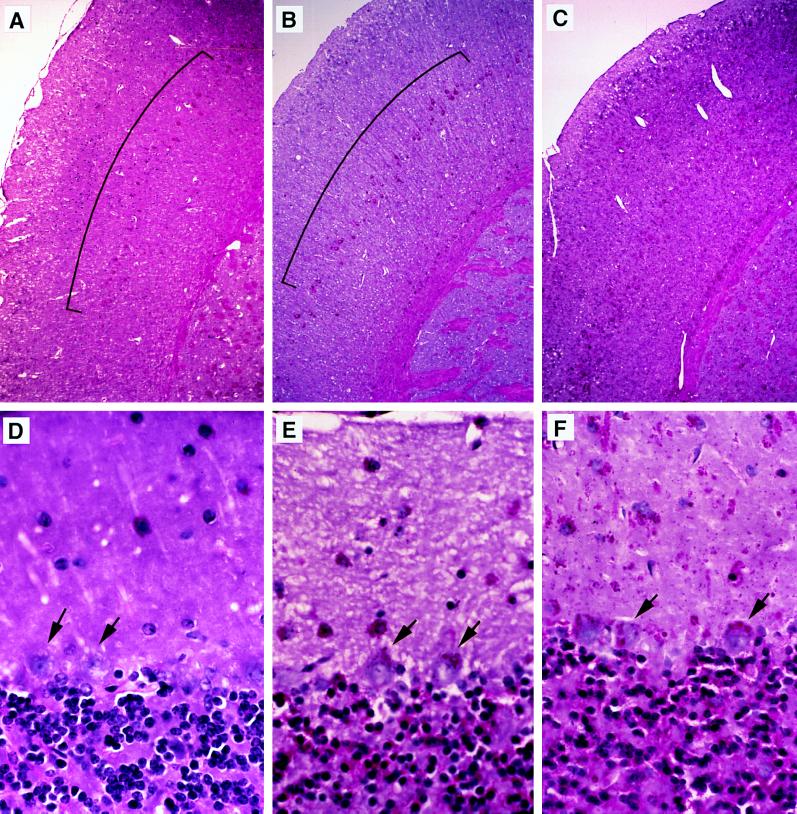
References
-
- Gravel R A, Clarke J T R, Kaback M M, Mahuran D, Sandhoff K, Suzuki K. In: The Metabolic and Molecular Basis of Inherited Disease. Scriver C R, Beaudet A L, Sly W S, Valle D, editors. New York: McGraw-Hill; 1995. pp. 2839–2879.
-
- Sandhoff K, Kolter T. Trends Cell Biol. 1996;6:98–103. - PubMed
-
- Taniike M, Yamanaka S, Proia R L, Langaman C, Bone-Turentine T, Suzuki K. Acta Neuropathol. 1995;89:296–304. - PubMed
-
- Sango K, Yamanaka S, Hoffmann A, Okuda Y, Grinberg A, Westphal H, McDonald M P, Crawley J N, Sandhoff K, Suzuki K, Proia R L. Nat Genet. 1995;11:170–176. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
