

Laminin 5 binds the NC-1 domain of type VII collagen
- PMID: 9245798
- PMCID: PMC2141627
- DOI: 10.1083/jcb.138.3.719
Laminin 5 binds the NC-1 domain of type VII collagen
Abstract
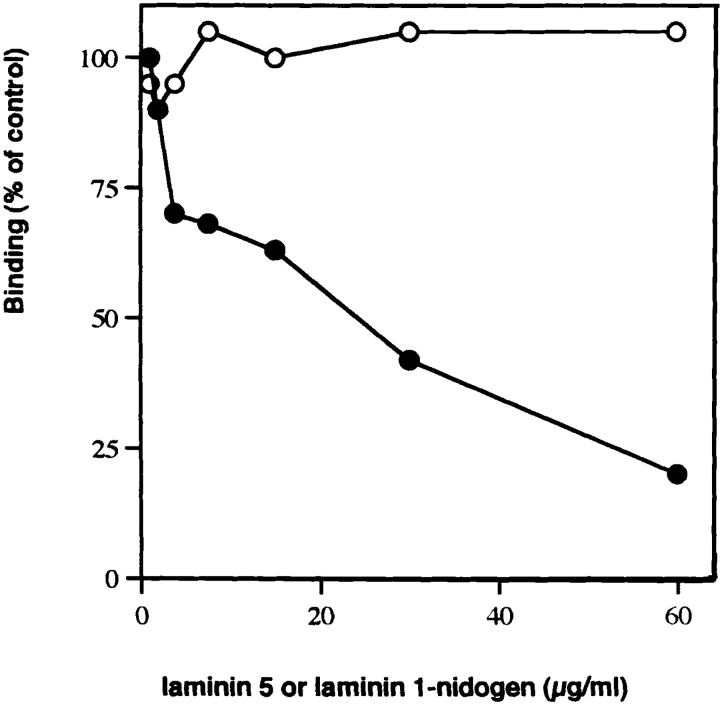
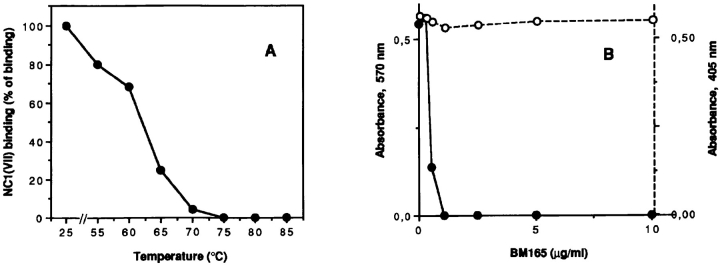
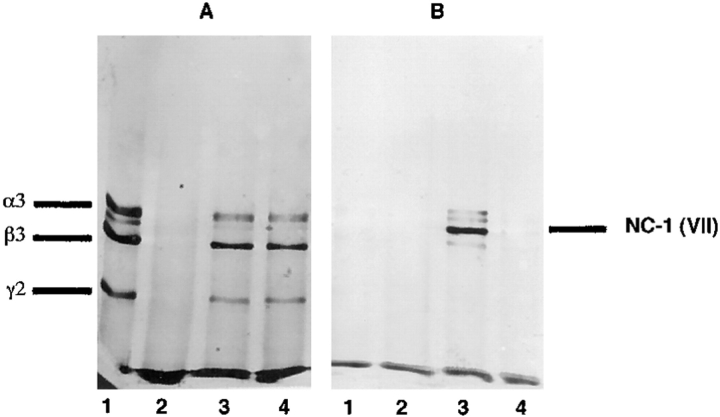
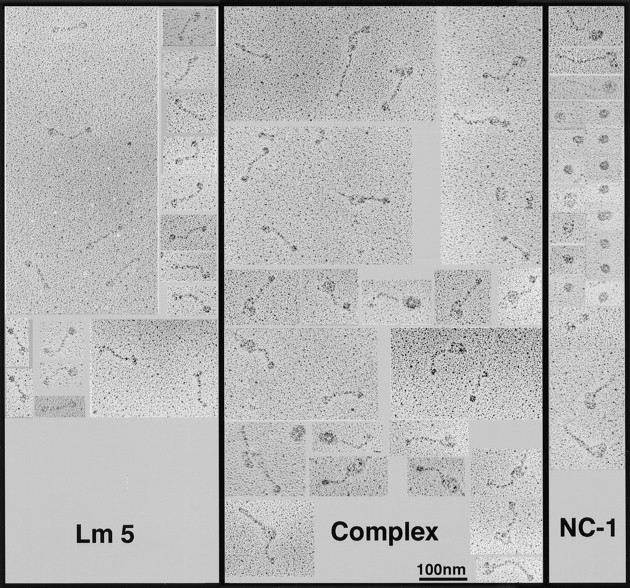
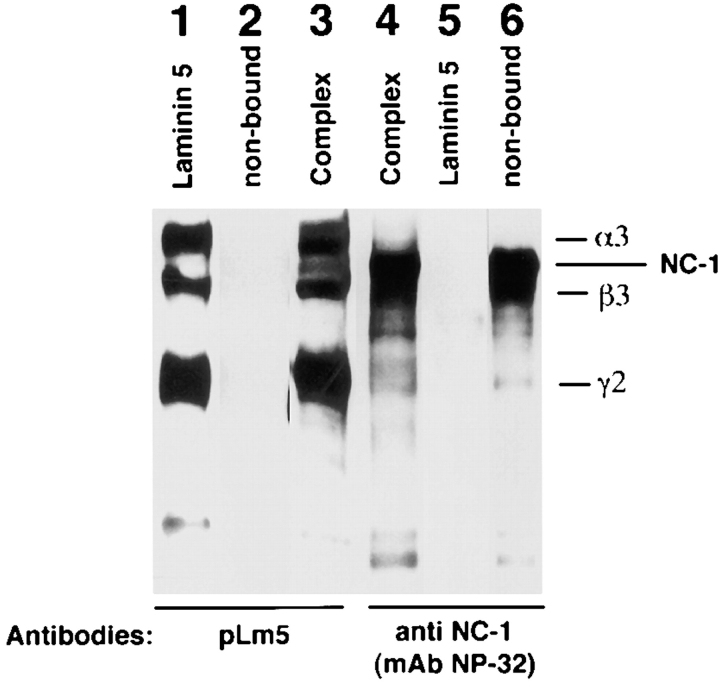
Mutational analyses of genes that encode components of the anchoring complex underlying the basolateral surface of external epithelia indicate that this structure is the major element providing for resistance to external friction. Ultrastructurally, laminin 5 (alpha3beta3gamma2; a component of the anchoring filament) appears as a thin filament bridging the hemidesmosome with the anchoring fibrils. Laminin 5 binds the cell surface through hemidesmosomal integrin alpha6beta4. However, the interaction of laminin 5 with the anchoring fibril (type VII collagen) has not been elucidated. In this study we demonstrate that monomeric laminin 5 binds the NH2-terminal NC-1 domain of type VII collagen. The binding is dependent upon the native conformation of both laminin 5 and type VII collagen NC-1. Laminin 6 (alpha3beta1gamma1) has no detectable affinity for type VII collagen NC-1, indicating that the binding is mediated by the beta3 and/or gamma2 chains of laminin 5. Approximately half of the laminin 5 solubilized from human amnion or skin is covalently complexed with laminins 6 or 7 (alpha3beta2gamma1). The adduction occurs between the NH2 terminus of laminin 5 and the branch point of the short arms of laminins 6 or 7. The results are consistent with the presumed orientation of laminin 5, having the COOH-terminal G domain apposed to the hemidesmosomal integrin, and the NH2-terminal domains within the lamina densa. The results also support a model predicting that monomeric laminin 5 constitutes the anchoring filaments and bridges integrin alpha6beta4 with type VII collagen, and the laminin 5-6/7 complexes are present within the interhemidesmosomal spaces bound at least by integrin alpha3beta1 where they may mediate basement membrane assembly or stability, but contribute less significantly to epithelial friction resistance.
Figures
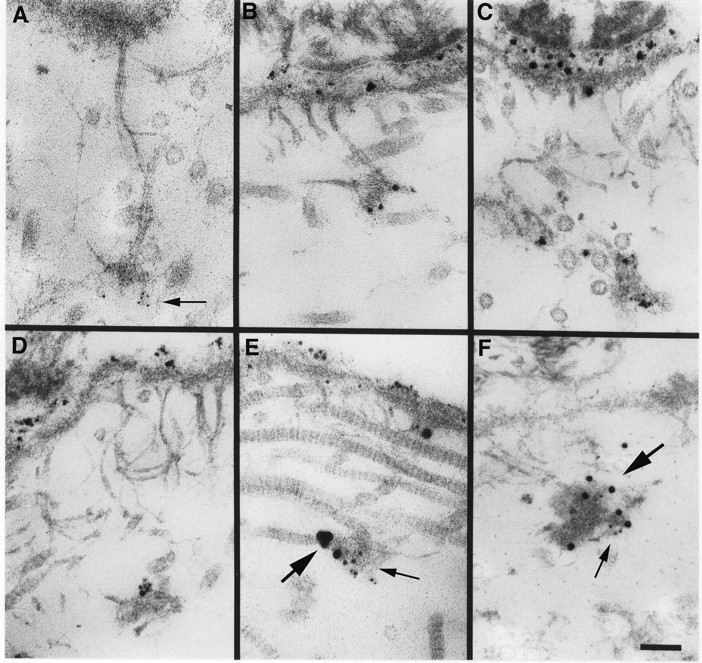
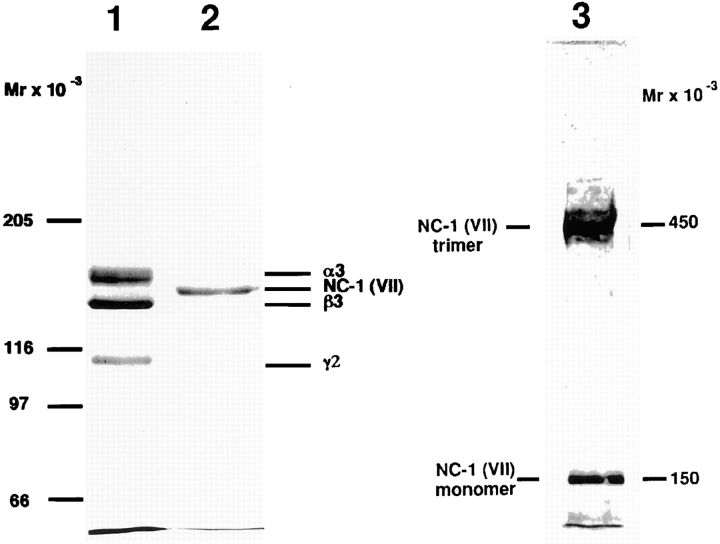
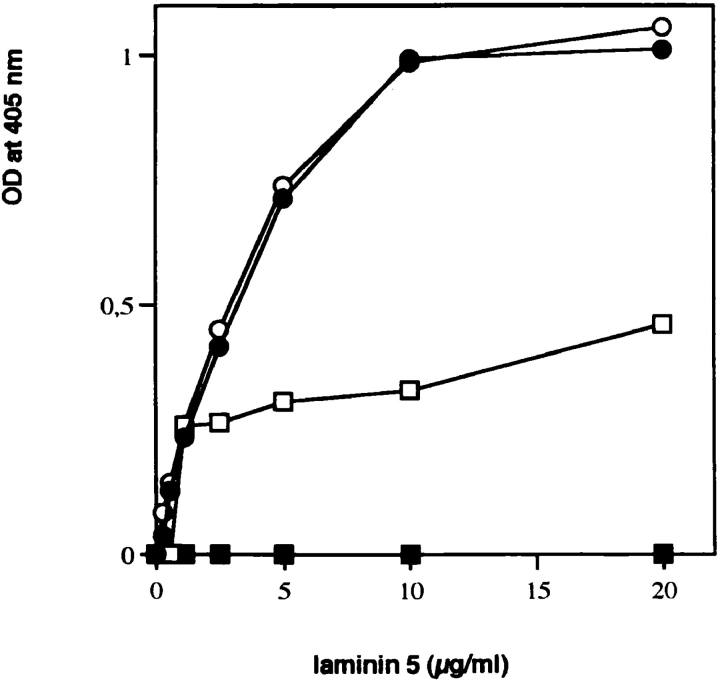
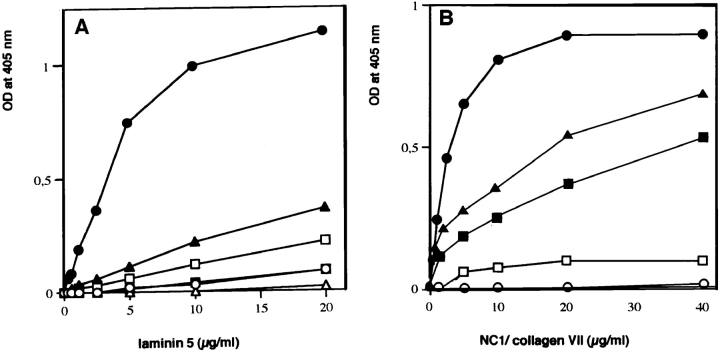

References
-
- Aubert-Foucher E, Font B, Eichenberger D, Goldschmidt D, Lethias C, van der Rest M. Purification and partial characterization of native type XIV collagen. J Biol Chem. 1992;267:15759–15764. - PubMed
-
- Aumailley M, Wiedemann H, Mann K, Timpl R. Binding of nidogen and the laminin-nidogen complex to basement membrane collagen type IV. Eur J Biochem. 1989;184:241–248. - PubMed
-
- Bachinger HP, Morris NP, Lunstrum GP, Keene DR, Rosenbaum LM, Compton LA, Burgeson RE. The relationship of the biophysical and biochemical characteristics of type VII collagen to the function of anchoring fibrils. J Biol Chem. 1990;265:10095–10101. - PubMed
-
- Bruckner-Tuderman L. Collagen VII and bullous disorders of the skin. Dermatology. 1994;189:16–20. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
