

Estrogen inhibits bone resorption by directly inducing apoptosis of the bone-resorbing osteoclasts
- PMID: 9254647
- PMCID: PMC2199029
- DOI: 10.1084/jem.186.4.489
Estrogen inhibits bone resorption by directly inducing apoptosis of the bone-resorbing osteoclasts
Abstract
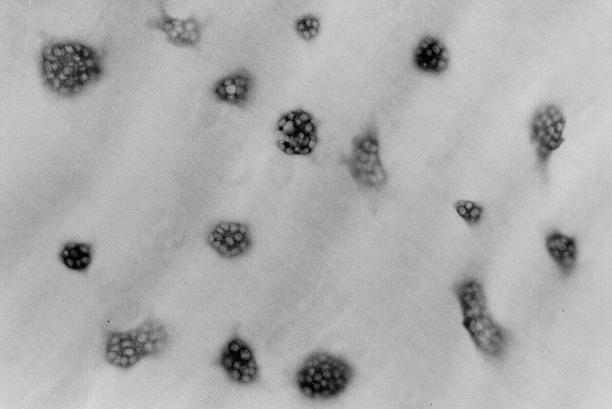
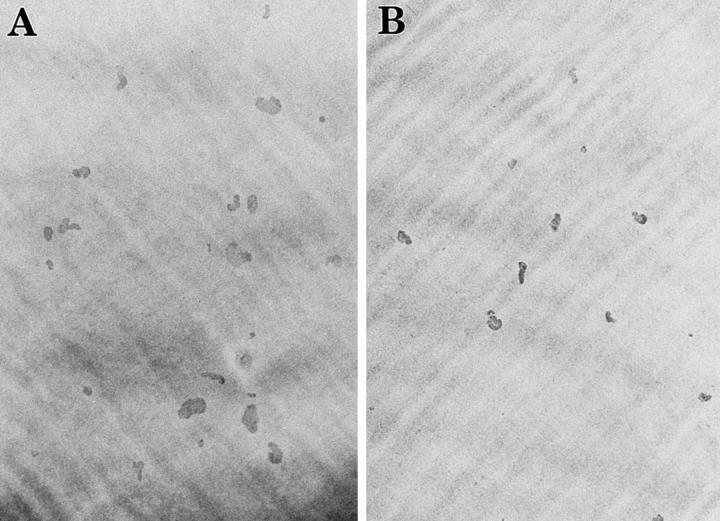
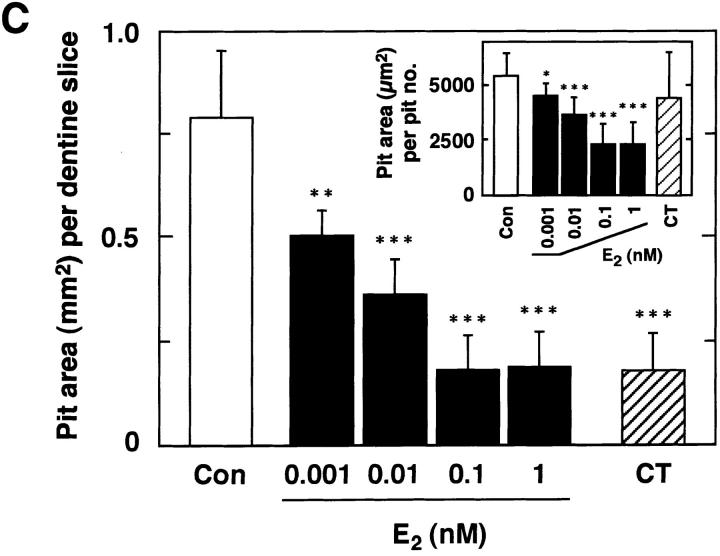
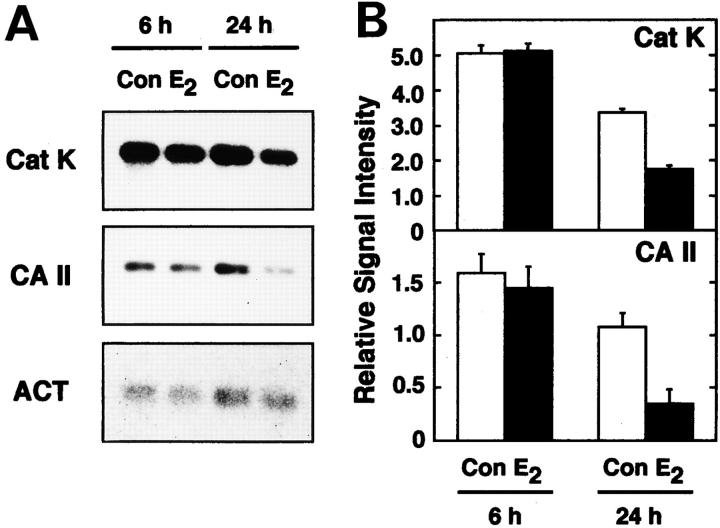
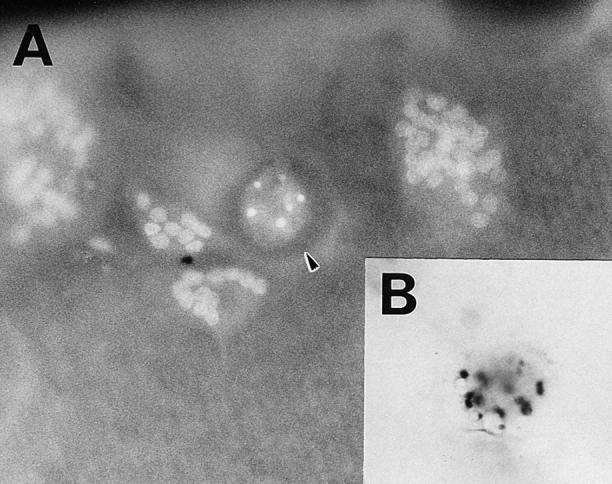
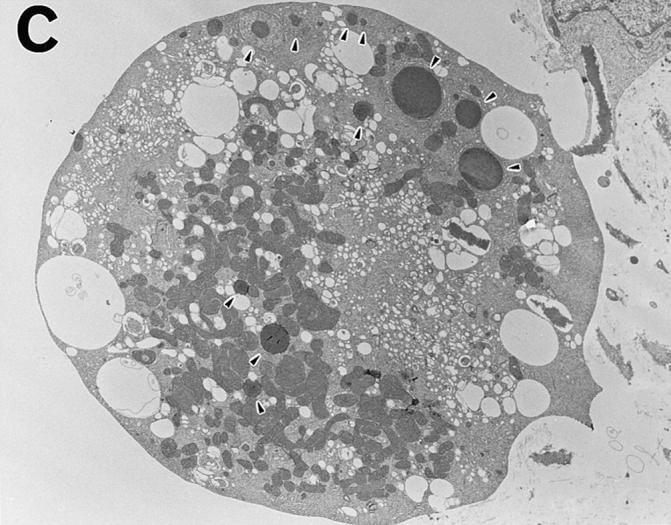
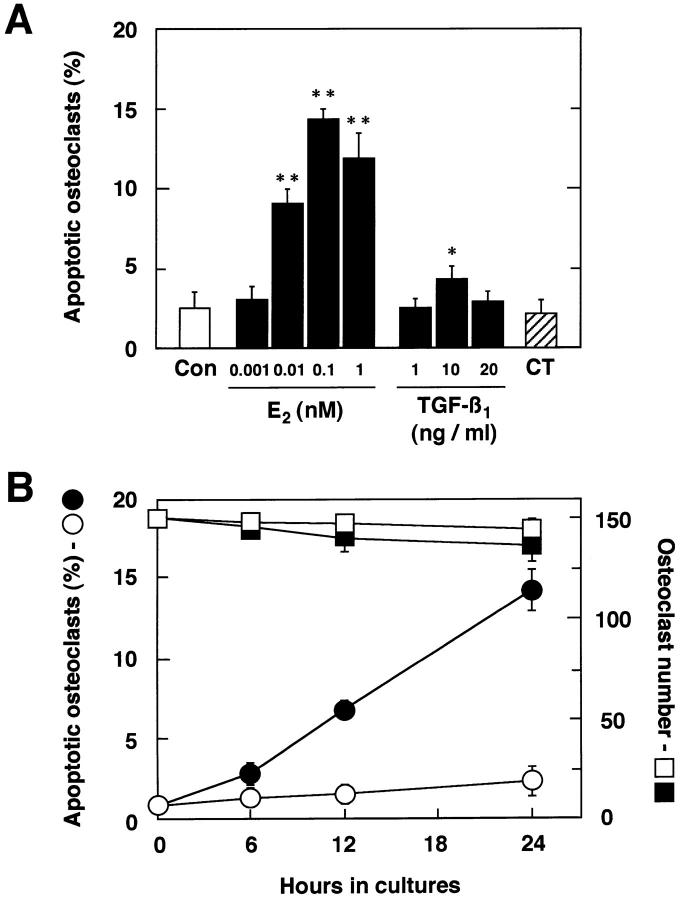
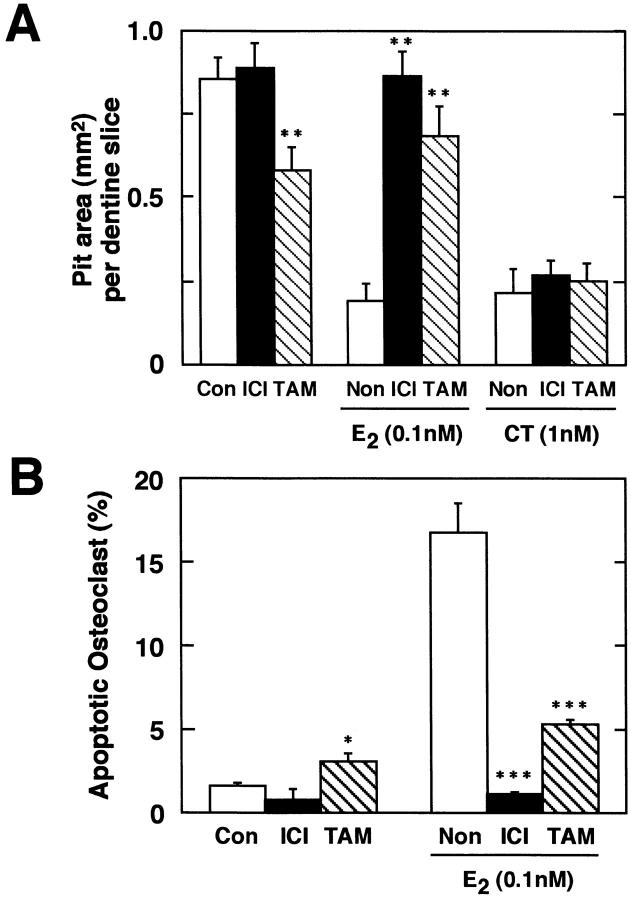
Estrogen deficiency causes bone loss, which can be prevented by estrogen replacement therapy. Using a recently developed technique for isolation of highly purified mammalian osteoclasts, we showed that 17 beta-estradiol (E2) was able to directly inhibit osteoclastic bone resorption. At concentrations effective for inhibiting bone resorption, E2 also directly induced osteoclast apoptosis in a dose- and time-dependent manner. ICI164,384 and tamoxifen, as pure and partial antagonists, respectively, completely or partially blocked the effect of E2 on both inhibition of osteoclastic bone resorption and induction of osteoclast apoptosis. These data suggest that the protective effects of estrogen against postmenopausal osteoporosis are mediated in part by the direct induction of apoptosis of the bone-resorbing osteoclasts by an estrogen receptor- mediated mechanism.
Figures
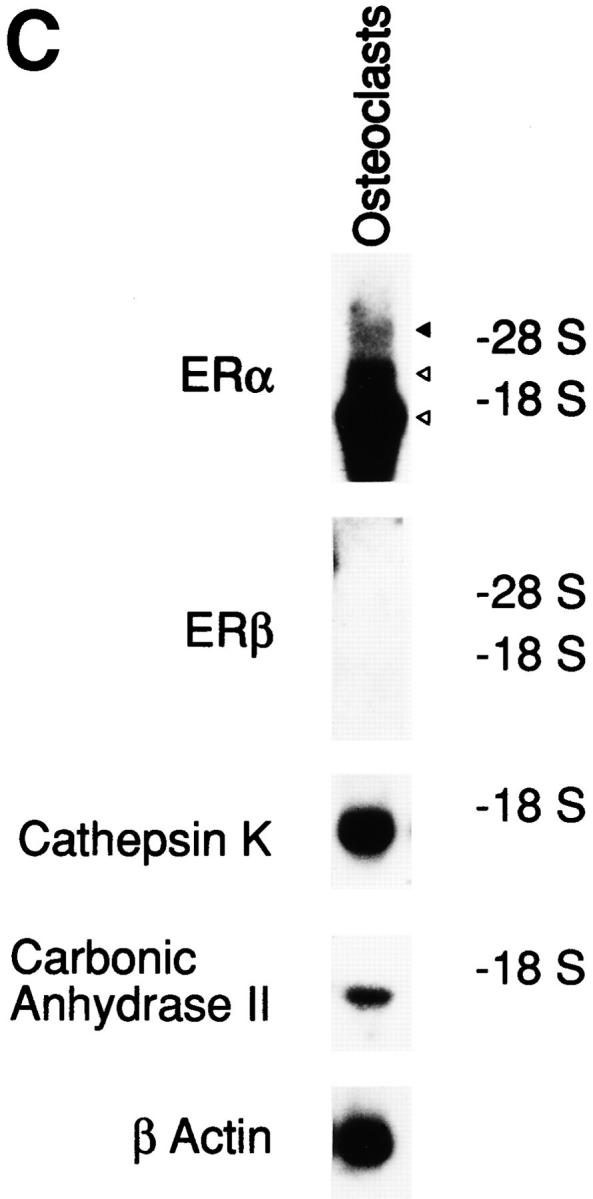
References
-
- Lindsay R, Hart DM, Aitken JM, MacDonald EB, Anderson JB, Clarke AC. Long-term prevention of postmenopausal osteoporosis by oestrogen. Evidence for an increased bone mass after delayed onset of oestrogen treatment. Lancet. 1976;1:1038–1041. - PubMed
-
- Ettinger B, Genant HK, Cann CE. Long-term estrogen replacement therapy prevents bone loss and fractures. Ann Intern Med. 1985;102:319–324. - PubMed
-
- Heaney RP, Recker RR, Saville PD. Menopausal changes in bone remodeling. J Lab Clin Med. 1978;92:964–970. - PubMed
-
- Selby PL, Peacock M, Barkworth SA, Brown WB, Taylor GA. Early effects of ethinyloestradiol and norethisterone treatment in post-menopausal women on bone resorption and calcium regulating hormones. Clin Sci (Lond) 1985;69:265–271. - PubMed
-
- Vaes G. Cellular biology and biochemical mechanism of bone resorption. A review of recent developments on the formation, activation, and mode of action of osteoclasts. Clin Orthop Relat Res. 1988;231:239–271. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
