

Developing neonatal rat sympathetic and sensory neurons differ in their regulation of 5-HT3 receptor expression
- PMID: 9254675
- PMCID: PMC6573128
- DOI: 10.1523/JNEUROSCI.17-17-06629.1997
Developing neonatal rat sympathetic and sensory neurons differ in their regulation of 5-HT3 receptor expression
Abstract
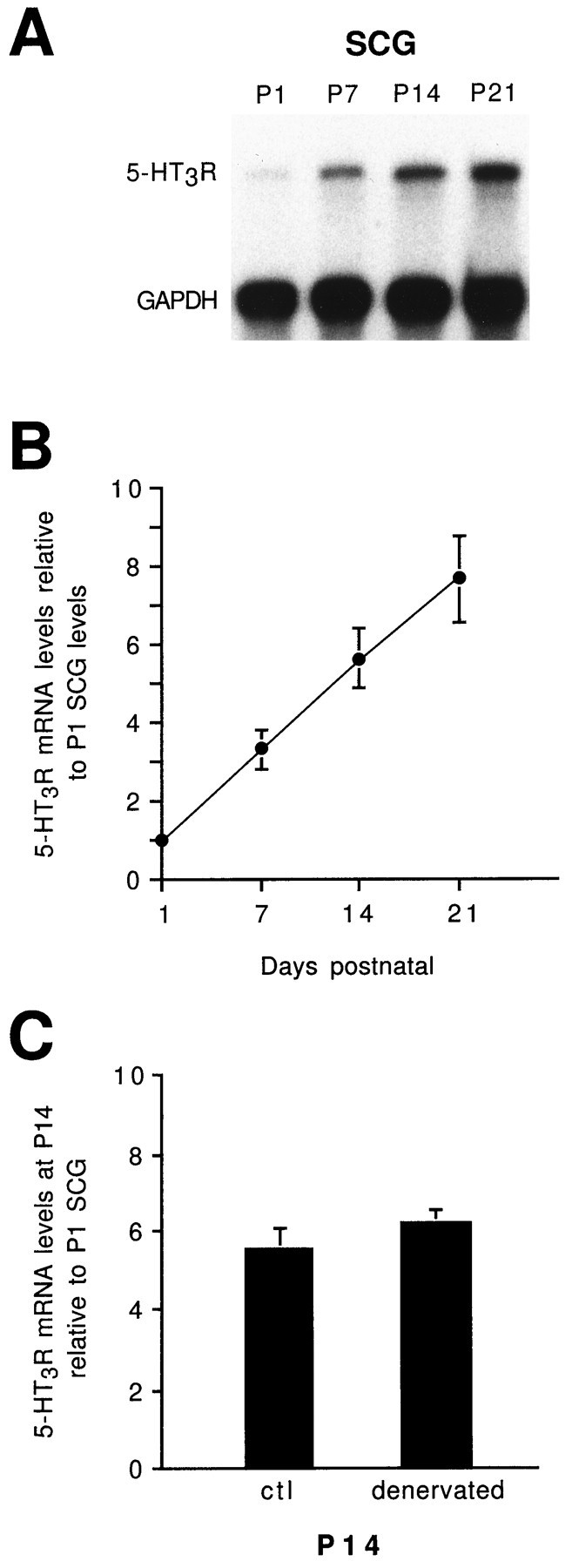
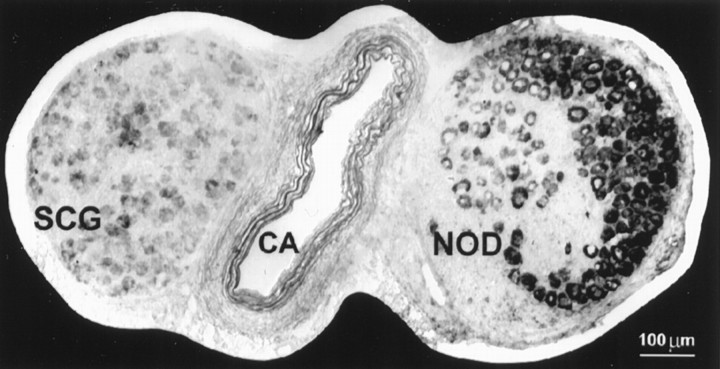
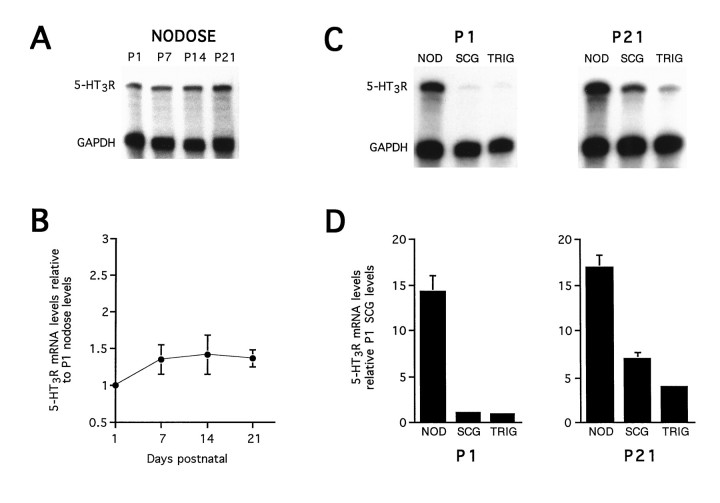
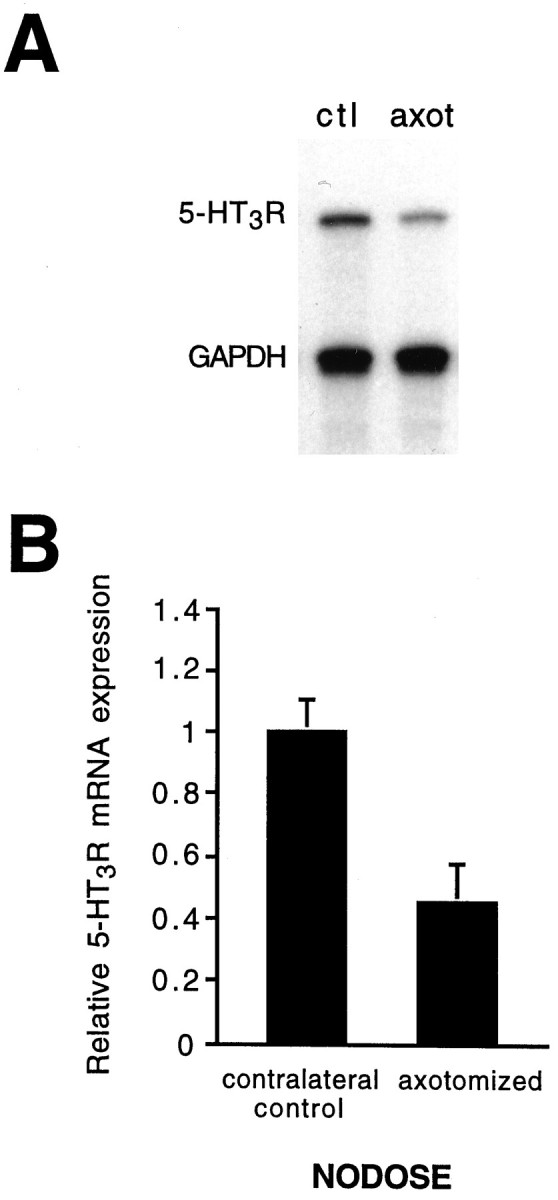
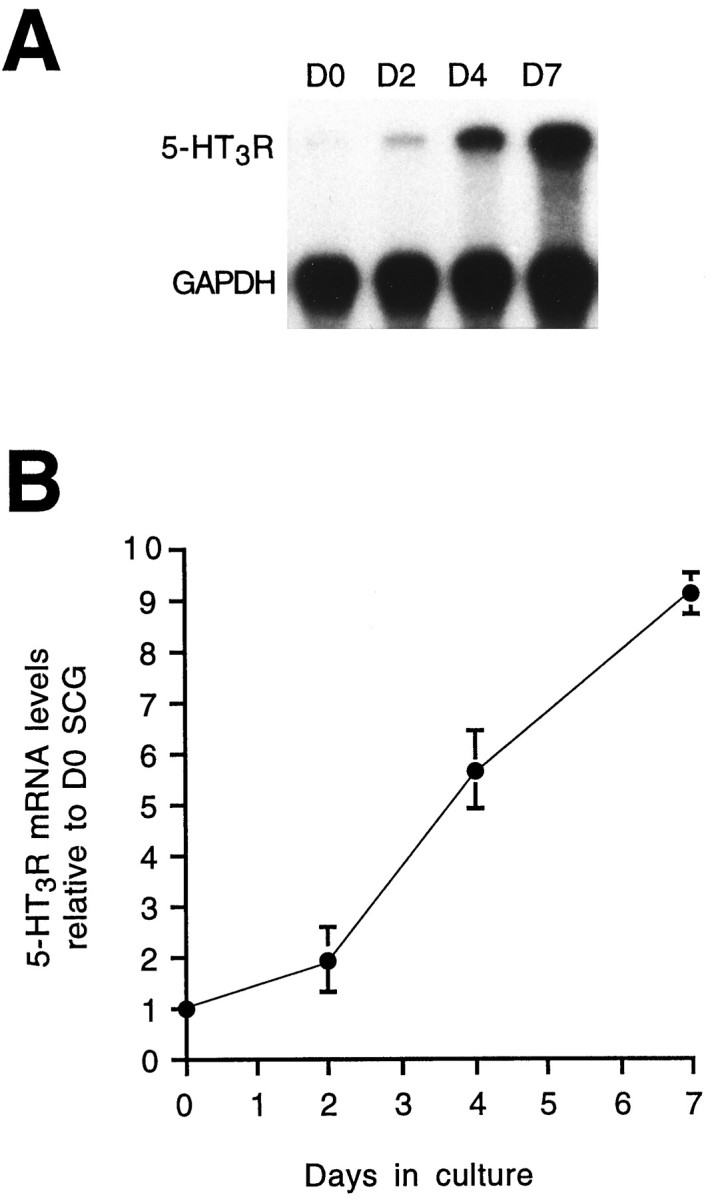
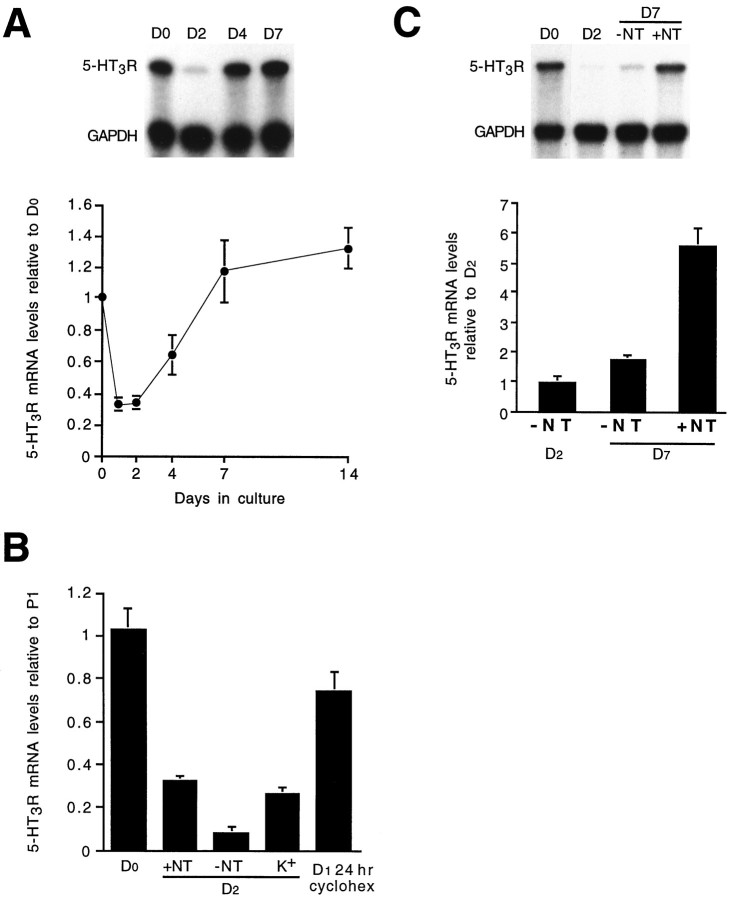
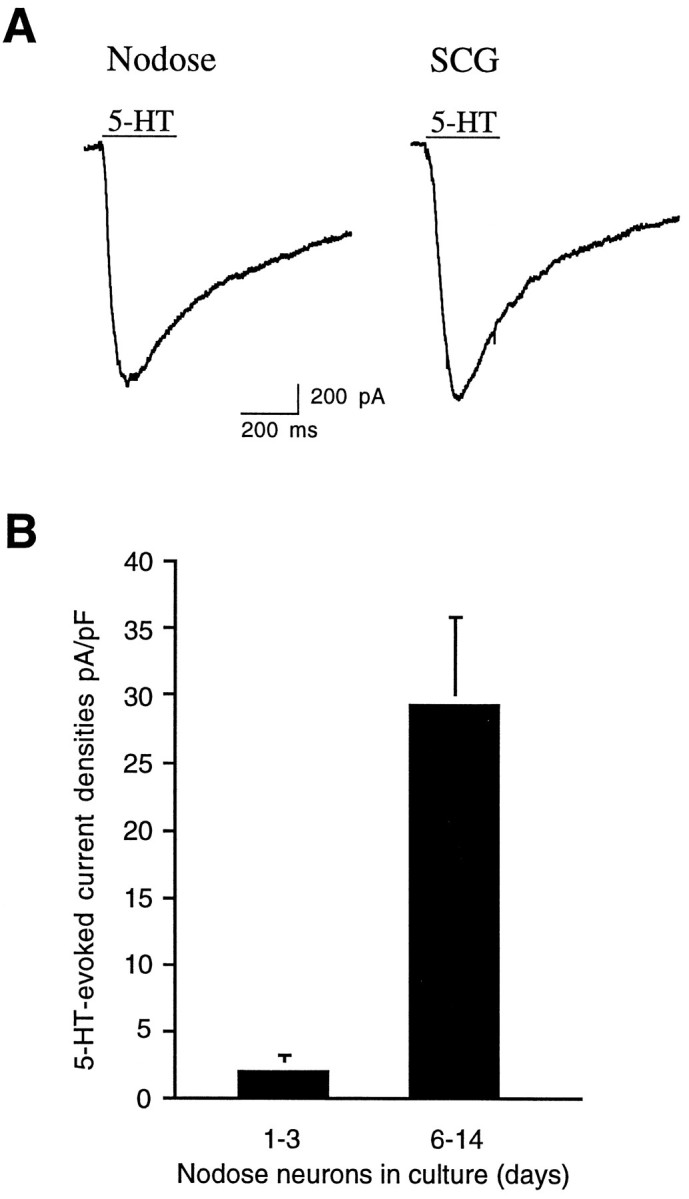
Serotonin 5-HT3 receptors (5-HT3Rs) are ligand-gated ion channels expressed by many peripheral neurons and are involved in several physiological processes. To learn more about the developmental regulation of 5-HT3R expression, we investigated rat sympathetic and vagal sensory neurons. We found that sympathetic and sensory neurons differ in their regulation of 5-HT3R expression during early postnatal life and as these neurons develop in culture. In SCG neurons 5-HT3R transcript levels are low at postnatal day 1 (P1) and increase 7.5-fold by P21; this increase occurs even after elimination of preganglionic innervation. In comparison, 5-HT3R mRNA levels in P1 nodose neurons are over 14-fold greater than in P1 SCG and change little by P21. We show that 5-HT3R transcript levels in nodose neurons depend on intact target innervation and drop by 60% after axotomy. When P1 SCG neurons develop in culture, we observed a significant increase in 5-HT3R expression: after 7 d in culture, transcript levels increase ninefold versus a threefold increase for neurons developing for 7 d in vivo. In contrast, 5-HT3R mRNA levels in cultured nodose neurons drop by 70% within 24 hr; however, this drop is transient. After 2 d, transcript levels begin to increase, and after 7 d, they are above initial values. We show that this delayed increase in 5-HT3R expression depends on neurotrophins. In both nodose and sympathetic neurons we found that the changes in 5-HT3R gene expression correlate directly with the appearance of 5-HT-evoked current densities.
Figures
References
-
- Andresen M, Kunze D. Nucleus tractus solitarius—gateway to neural circulatory control. Annu Rev Physiol. 1994;56:93–116. - PubMed
-
- Apud JA. The 5-HT3 receptor in mammalian brain: a new target for the development of psychotropic drugs? Neuropsychopharmacology. 1993;8:117–130. - PubMed
-
- Barthel L, Raymond P. Subcellular localization of α-tubulin and opsin mRNA in the goldfish retina using digoxigenin-labeled cRNA probes detected by alkaline phosphatase and HRP histochemistry. J Neurosci Methods. 1993;50:145–152. - PubMed
-
- Black IB. Regulation of autonomic development. Annu Rev Neurosci. 1978;1:183–214. - PubMed
-
- Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidium thiocyanate-phenol-chloroform extraction. Anal Biochem. 1987;162:156–159. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical