

Spatial and temporal expression of the period and timeless genes in the developing nervous system of Drosophila: newly identified pacemaker candidates and novel features of clock gene product cycling
- PMID: 9254686
- PMCID: PMC6573141
- DOI: 10.1523/JNEUROSCI.17-17-06745.1997
Spatial and temporal expression of the period and timeless genes in the developing nervous system of Drosophila: newly identified pacemaker candidates and novel features of clock gene product cycling
Abstract
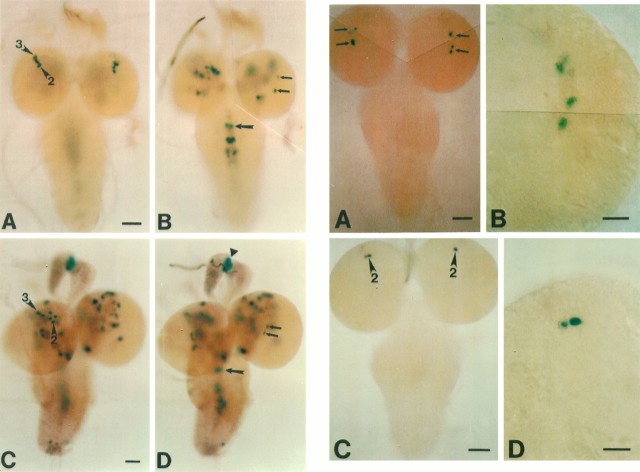
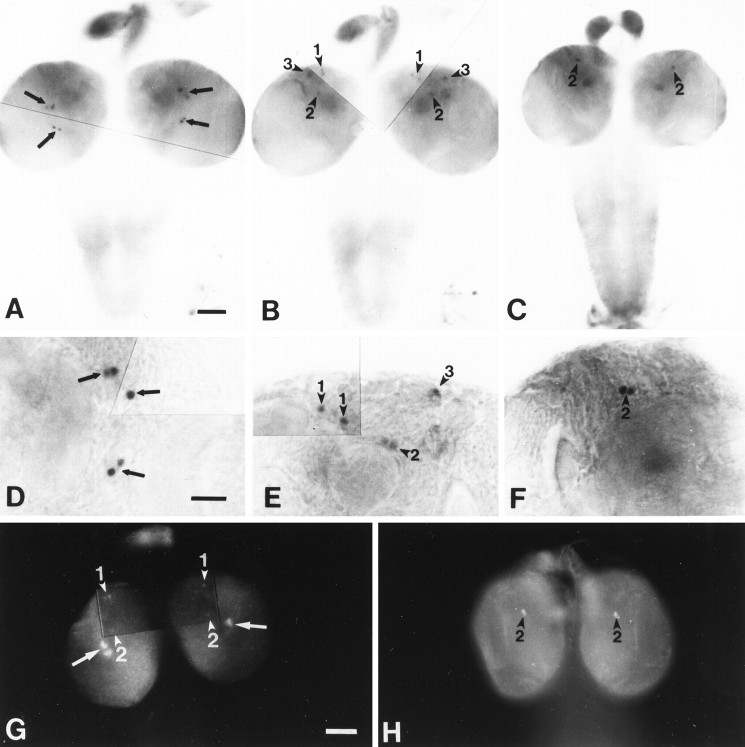
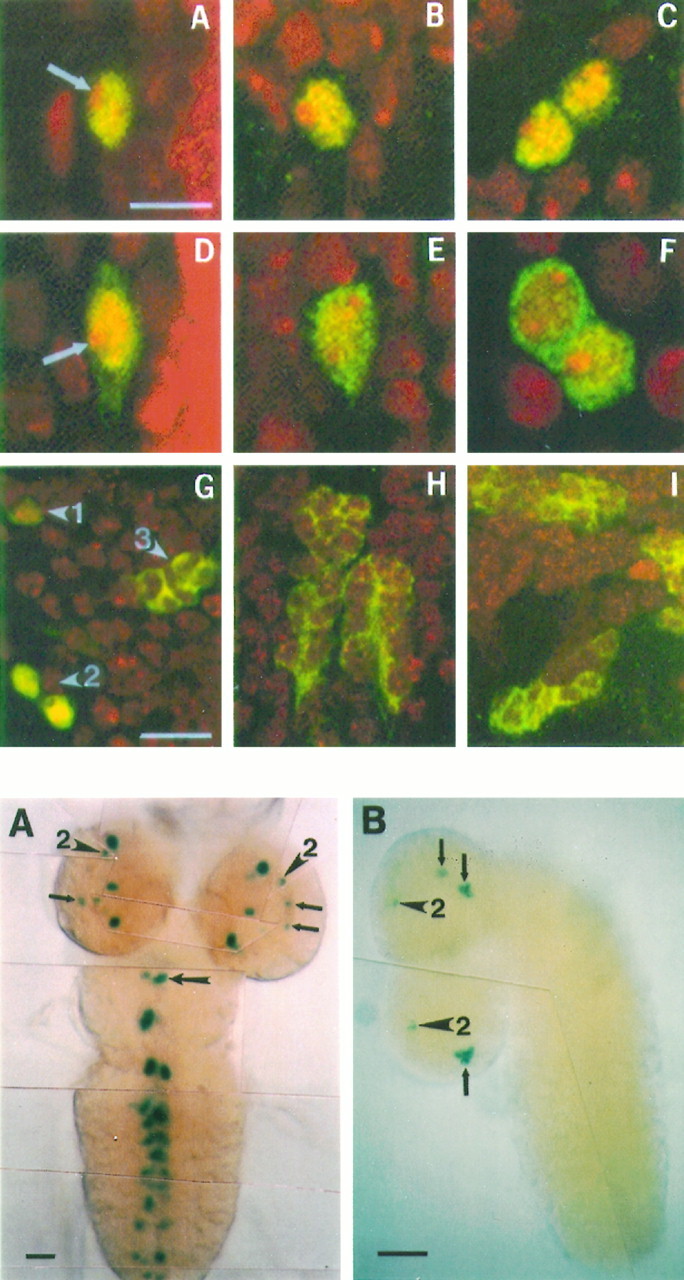
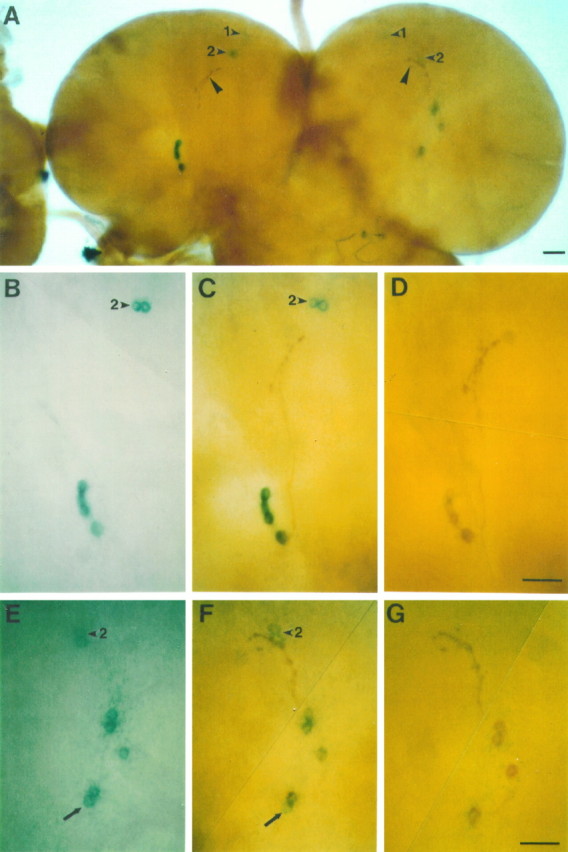
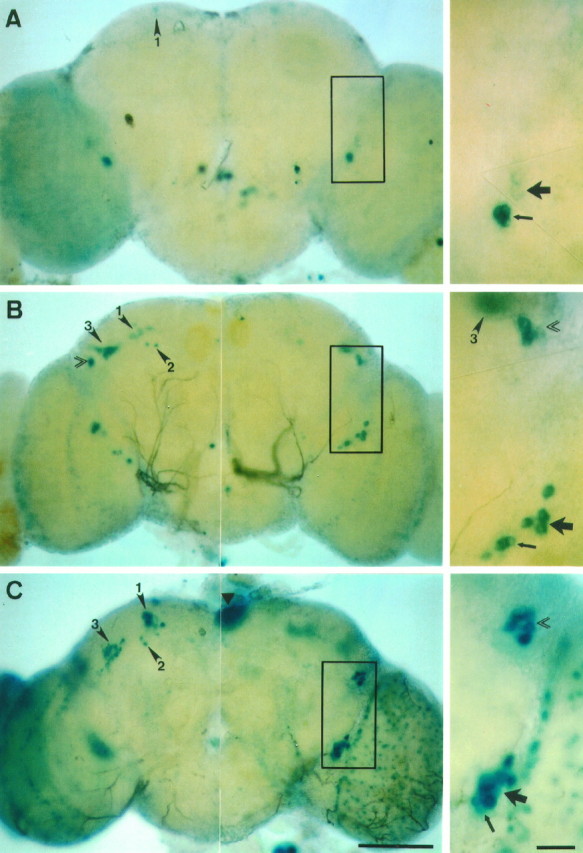
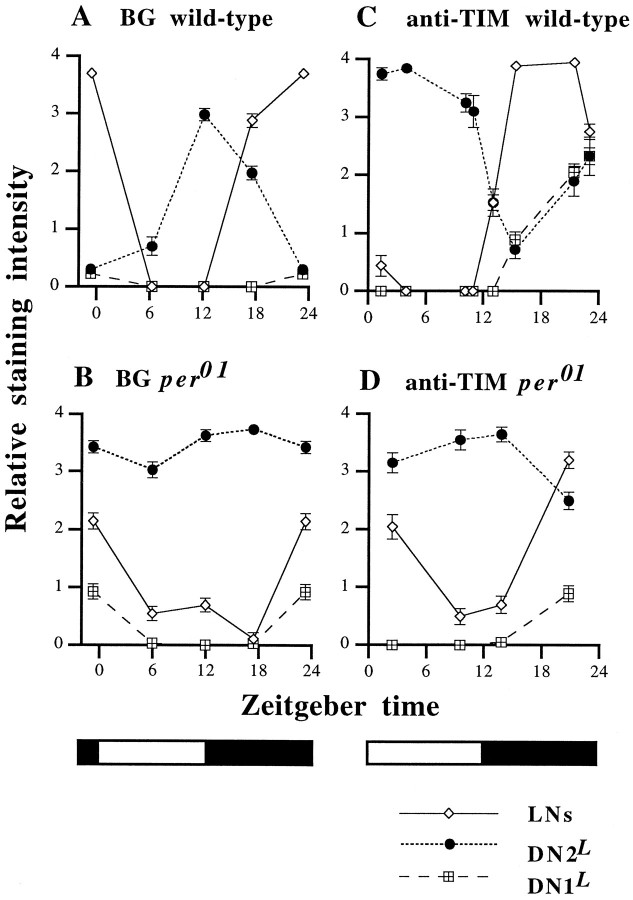
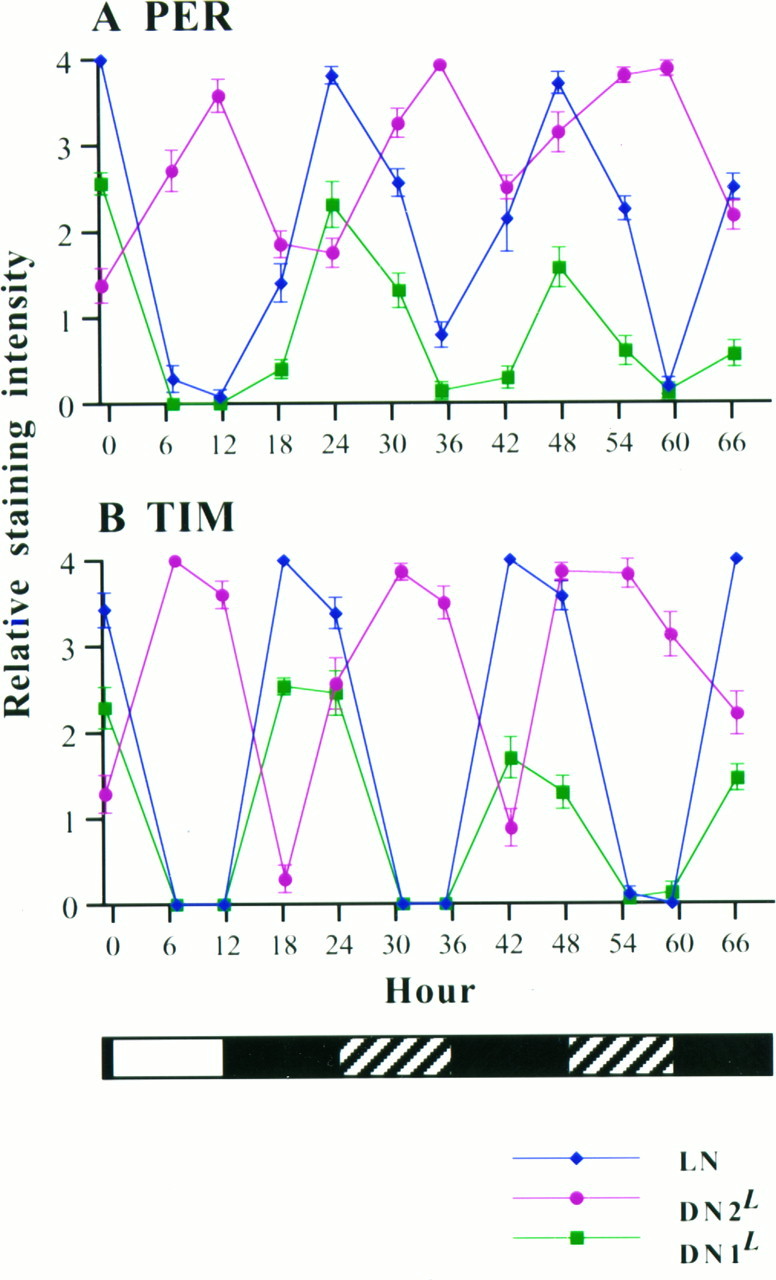
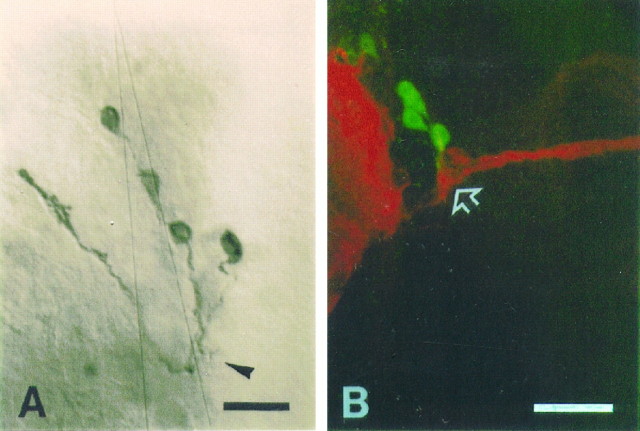
The circadian timekeeping system of Drosophila functions from the first larval instar (L1) onward but is not known to require the expression of clock genes in larvae. We show that period (per) and timeless (tim) are rhythmically expressed in several groups of neurons in the larval CNS both in light/dark cycles and in constant dark conditions. Among the clock gene-expressing cells there is a subset of the putative pacemaker neurons, the "lateral neurons" (LNs), that have been analyzed mainly in adult flies. Like the adult LNs, the larval ones are also immunoreactive to a peptide called pigment-dispersing hormone. Their putative dendritic trees were found to be in close proximity to the terminals of the larval optic nerve Bolwig's nerve, possibly receiving photic input from the larval eyes. The LNs are the only larval cells that maintain a strong cycling in PER from L1 onward, throughout metamorphosis and into adulthood. Therefore, they are the best candidates for being pacemaker neurons responsible for the larval "time memory" (inferred from previous experiments). In addition to the LNs, a subset of the larval dorsal neurons (DNLs) expresses per and tim. Intriguingly, two neurons of this DNL group cycle in PER and TIM immunoreactivity almost in antiphase to the other DNLs and to the LNs. Thus, the temporal expression of per and tim are regulated differentially in different cells. Furthermore, the light sensitivity associated with levels of the TIM protein is different from that in the heads of adult Drosophila.
Figures
References
-
- Amrein H, Axel R. Genes expressed in neurons of adult male Drosophila. Cell. 1997;88:459–469. - PubMed
-
- Bargiello TA, Saez L, Baylies MK, Gasic G, Young MW, Spray DC. The Drosophila clock gene per affects intercellular junctional communication. Nature. 1987;328:686–691. - PubMed
-
- Bier E, Ackerman L, Barbel S, Jan L, Jan YN. Identification and characterization of a neuron-specific nuclear antigen in Drosophila. Science. 1988;240:913–916. - PubMed
-
- Bodenstein D. The postembryonic development of Drosophila. In: Demerec M, editor. Biology of Drosophila. Wiley; New York: 1950. pp. 275–367.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases