

doi: 10.1073/pnas.94.17.9202.
The catalytic subunit of yeast telomerase
Affiliations
- PMID: 9256460
- PMCID: PMC23115
- DOI: 10.1073/pnas.94.17.9202
Item in Clipboard
The catalytic subunit of yeast telomerase
Proc Natl Acad Sci U S A.
.
Abstract
Telomerase is an RNA-directed DNA polymerase, composed of RNA and protein subunits, that replicates the telomere ends of linear eukaryotic chromosomes. Using a genetic strategy described here, we identify the product of the EST2 gene, Est2p, as a subunit of telomerase in the yeast Saccharomyces cerevisiae. Est2p is required for enzyme catalysis, as mutations in EST2 were found to result in the absence of telomerase activity. Immunochemical experiments show that Est2p is an integral subunit of the telomerase enzyme. Critical catalytic residues present in RNA-directed DNA polymerases are conserved in Est2p; mutation of one such residue abolishes telomerase activity, suggesting a direct catalytic role for Est2p.
Figures
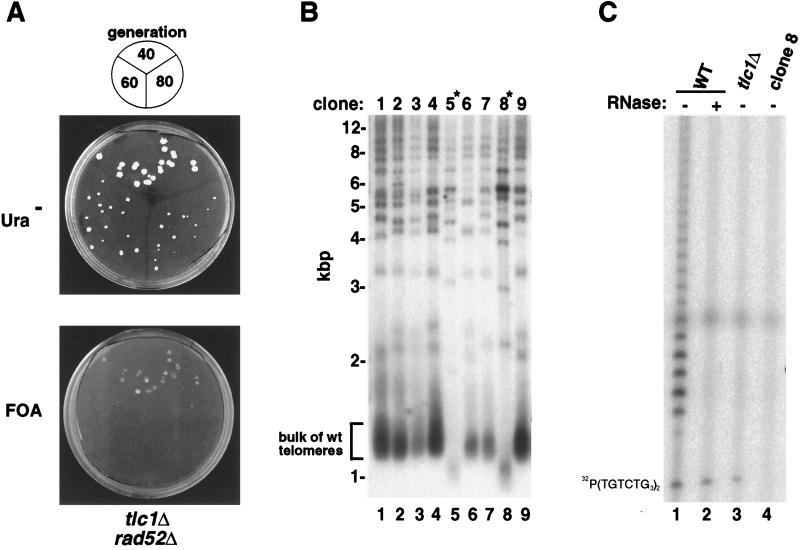
A genetic screen identifies genes required for telomerase activity. (A) Demonstration of the feasibility of the rad52 synthetic lethal screen. tlc1Δ rad52Δ haploid yeast harboring a plasmid expressing Rad52 protein were aged for 40, 60, or 80 generations and replica-plated on medium lacking uracil (Ura−) to retain the RAD52 plasmid (Upper) or on FOA to evict the RAD52 plasmid (Lower). (B) Identification of mutant clones with short telomeres (∗). Telomere hybridization analysis of XhoI-digested genomic DNA isolated from a subset of mutant clones that depend upon the RAD52-expressing plasmid for viability. The bulk of the wild-type (wt) telomeres appear as heterogeneous size fragments at ≈1.3 kbp. (C) Identification of a mutant clone lacking telomerase activity. Extracts were isolated from wild-type (WT) parental strain Y0025, from a tlc1Δ strain, and from mutant clone 8 and were incubated with (lane 2) or without (lanes 1, 3, and 4) RNase prior to assaying telomerase activity. Position of the γ-32P-labeled primer is shown on the left.
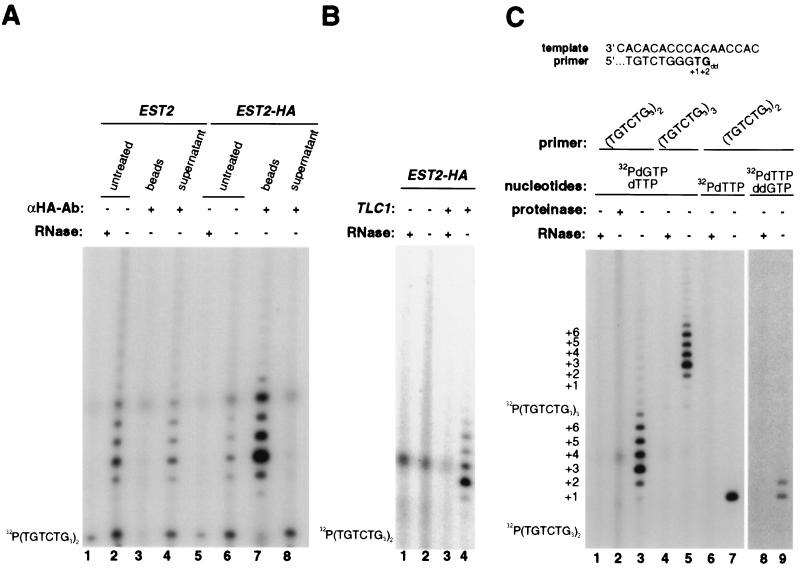
Est2p is a component of telomerase. (A) Immunoprecipitation and immunodepletion of telomerase activity by anti-HA antibody from EST2-HA yeast. Telomerase activity was assayed from lysates isolated from yeast strains expressing wild-type (EST2, lanes 1–4) or HA-tagged (EST2-HA, lanes 5–8) Est2p. The extracts were left untreated (lanes 2 and 6), treated with RNase (lanes 1 and 5), or incubated with protein A-agarose beads preloaded with anti-HA antibody and then centrifuged to separate the beads (lanes 3 and 7) from the supernatant (lanes 4 and 8). Position of the γ-32P-labeled primer is shown on the left. (B) Telomerase activity of Est2p-HA immunoprecipitates requires functional Tlc1. Telomerase activity was assayed on anti-HA immunoprecipitates from extracts of EST2-HA yeast with either a mutant tlc1Δ allele (lanes 1 and 2) or a wild-type TLC1 allele (lanes 3 and 4). RNase was added to duplicate reactions as a negative control (lanes 1 and 3). Position of the γ-32P-labeled primer is shown on the left. (C) The telomerase activity of Est2p-HA immunoprecipitates uses Tlc1 as a template. (Upper) Predicted alignment of the primer and the telomerase template, showing expected elongation products generated in the presence of [α-32P]dTTP alone or in conjunction with ddGTP. (Lower) Anti-HA immunoprecipitates from extracts of yeast expressing Est2p-HA were preincubated with either proteinase K (lane 1) or RNase (lanes 2, 4, 6, and 8) or were left untreated (lanes 3, 5, 7, and 9) prior to assaying telomerase. Telomerase activity was assayed with primer (TGTCTG3)2 (lanes 1–3 and 6–9) or (TGTCTG3)3 (lanes 4 and 5) in the presence of the deoxynucleoside triphosphate(s) [α-32P]dGTP and dTTP (lanes 1–5), or [α-32P]dTTP alone (lanes 6 and 7), or [α-32P]dTTP and ddGTP (lanes 8 and 9). A longer exposure is shown for lanes 8 and 9. The positions of the γ-32P-labeled primers and the elongation products from +1 to +6 are shown on the left.
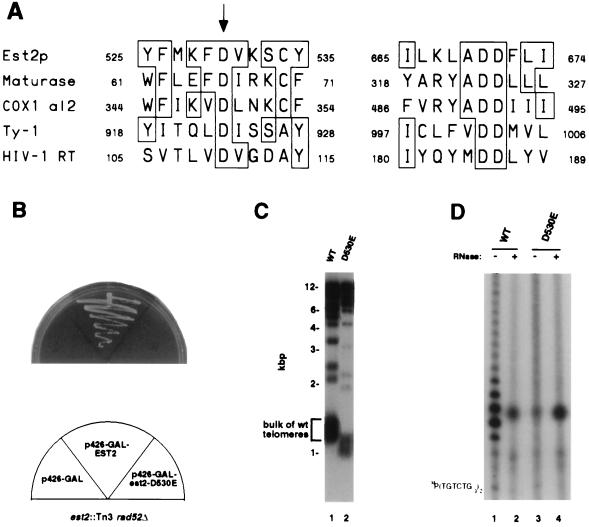
Reverse transcriptase homology of Est2p is essential for telomerase function. (A) Homology between reverse transcriptase catalytic residues and Est2p. Alignment of regions of Est2p containing the conserved reverse transcriptase motifs A and C (36), or 3 and 5 (37) with the same regions of Arabidopsis maturase, S. cerevisiae COX1 group II intron 2 and retrotransposon Ty-1, and HIV-1 reverse transcriptase (36, 37). Boxes denote residues conserved with Est2p. The three aspartic acid residues conserved among all the proteins shown are completely invariant among all RNA-dependent DNA polymerases (36, 37). The arrow denotes the Asp-530 residue that was mutagenized. (B) Failure of the D530E mutant to rescue inviability of est2 rad52 mutants. est2 rad52 mutant yeast were transformed with the negative control plasmid p426-GAL (left), with a positive control p426-GAL expressing wild-type EST2 (center), and with a p426-GAL plasmid expressing the D530E mutant (right). Loss of viability in the unrescued yeast is immediate, not delayed, because the est2 rad52 mutants have already been passaged ≈30 generations in culture. (C) Telomere shortening in est2 yeast expressing D530E mutant but not wild-type EST2. Telomere hybridization analysis of XhoI-digested genomic DNA isolated from an EST2 control (lane 1) or from est2 mutants expressing the D530E mutant (lane 2). (D) Lack of telomerase activity in yeast expressing the D530E mutant. Telomerase activity was assayed in extracts from an EST2 wild-type control (lanes 1 and 2) or from yeast expressing the D530E mutant (lanes 3 and 4). Extracts were pretreated with RNase (lanes 1 and 3) or left untreated (lanes 2 and 4). Position of the γ-32P-labeled primer is shown on the left.
References
-
- Blackburn E H. Annu Rev Biochem. 1984;53:163–194. - PubMed
-
- Zakian V A. Science. 1995;270:1601–1607. - PubMed
-
- Greider C W, Blackburn E H. Cell. 1985;43:405–413. - PubMed
-
- Greider C W. In: Telomerase Biochemistry and Regulation. Blackburn E H, Greider C W, editors. Plainview, NY: Cold Spring Harbor Lab. Press; 1995. pp. 35–68.
-
- Greider C W, Blackburn E H. Nature (London) 1989;337:331–337. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
