

Acid-induced dissociation of VacA, the Helicobacter pylori vacuolating cytotoxin, reveals its pattern of assembly
- PMID: 9265644
- PMCID: PMC2138037
- DOI: 10.1083/jcb.138.4.759
Acid-induced dissociation of VacA, the Helicobacter pylori vacuolating cytotoxin, reveals its pattern of assembly
Abstract
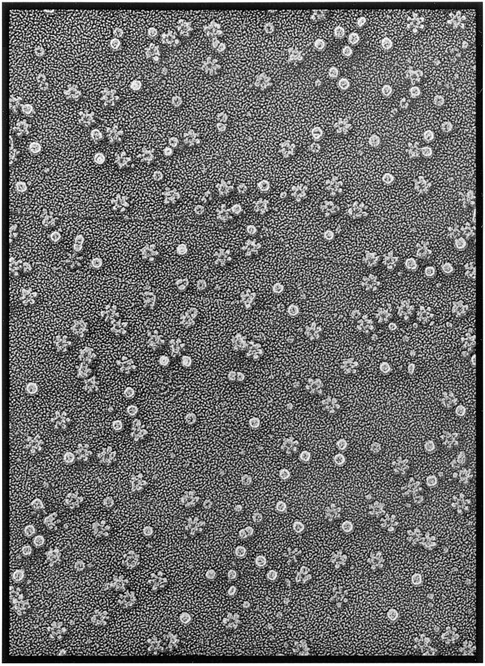
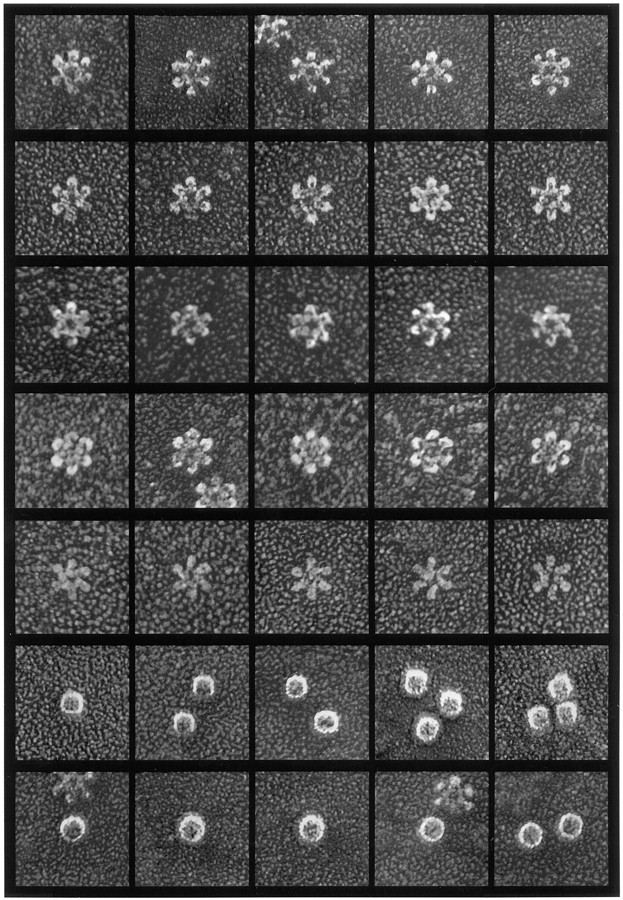
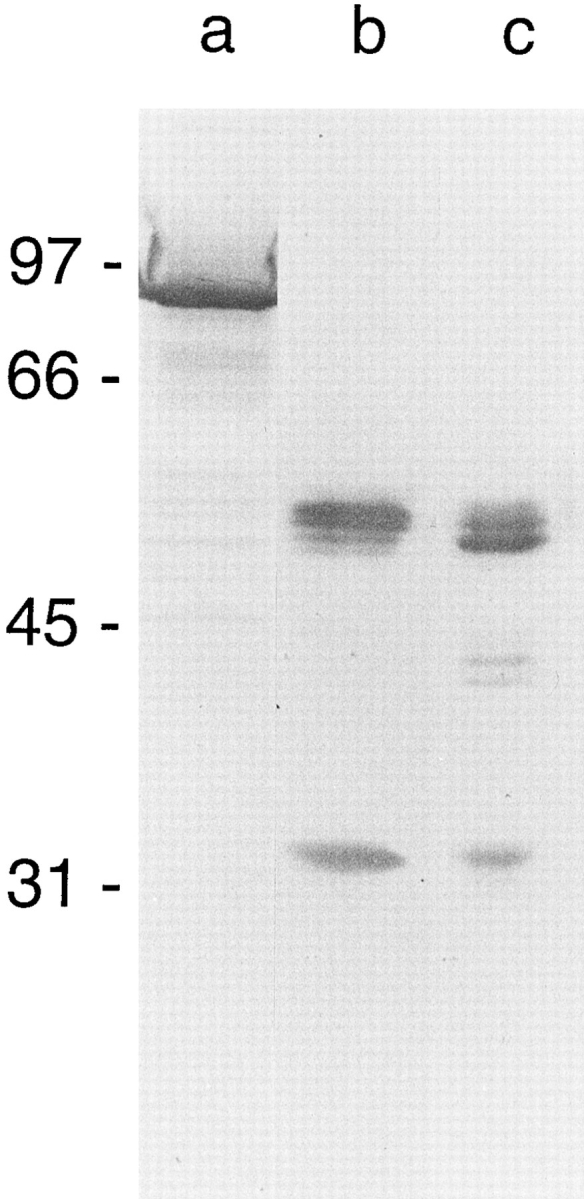
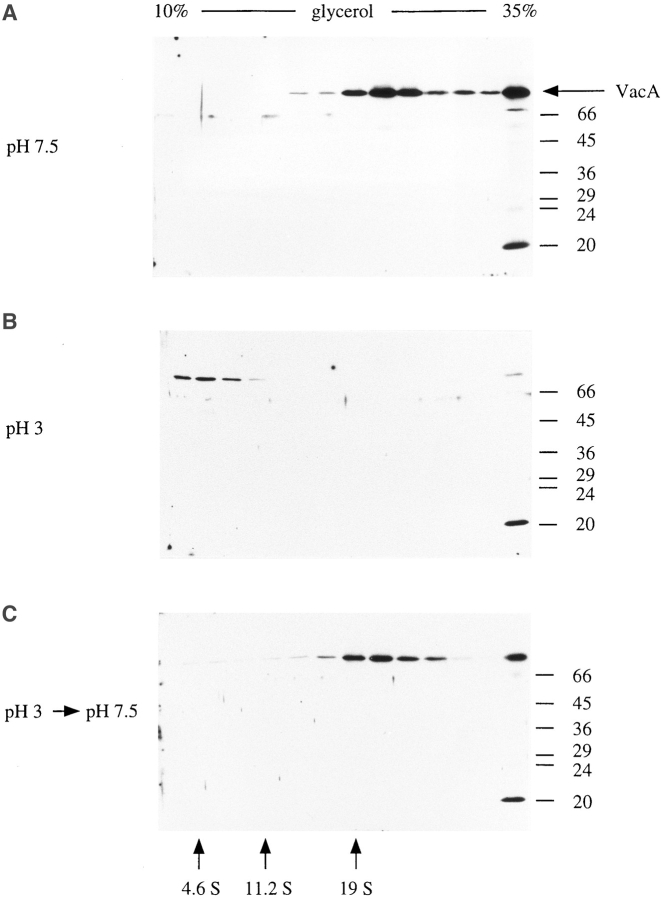
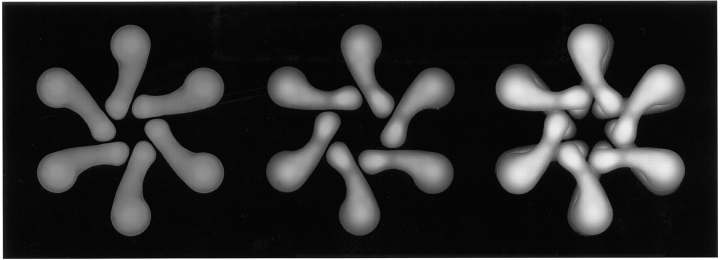
In this study, we describe the ultrastructural changes associated with acid activation of Helicobacter pylori vacuolating cytotoxin (VacA). Purified VacA molecules imaged by deep-etch electron microscopy form approximately 30-nm hexagonal "flowers," each composed of an approximately 15-nm central ring surrounded by six approximately 6-nm globular "petals." Upon exposure to acidic pH, these oligomeric flowers dissociate into collections of up to 12 teardrop-shaped subunits, each measuring approximately 6 x 14 nm. Correspondingly, glycerol density gradient centrifugation shows that at neutral pH VacA sediments at approximately 22 S, whereas at acidic pH it dissociates and sediments at approximately 5 S. Immunoblot and EM analysis of the 5-S material demonstrates that it represents approximately 90-kD monomers with 6 x 14-nm "teardrop" morphology. These data indicate that the intact VacA oligomer consists of 12 approximately 90-kD subunits assembled into two interlocked six-membered arrays, overlap of which gives rise to the flower-like appearance. Support for this interpretation comes from EM identification of small numbers of relatively "flat" oligomers composed of six teardrop-shaped subunits, interpreted to be halves of the complete flower. These flat forms adsorb to mica in two different orientations, corresponding to hexameric surfaces that are either exposed or sandwiched inside the dodecamer, respectively. This view of VacA structure differs from a previous model in which the flowers were interpreted to be single layers of six monomers and the flat forms were thought to be proteolysed flowers. Since acidification has been shown to potentiate the cytotoxic effects of VacA, the present results suggest that physical disassembly of the VacA oligomer is an important feature of its activation.
Figures






References
-
- Atherton JC, Cao P, Peek RM, Jr, Tummuru MKR, Blaser MJ, Cover TL. Mosaicism in vacuolating cytotoxin alleles of Helicobacter pylori: association of specific vacA types with cytotoxin production and peptic ulceration. J Biol Chem. 1995;270:17771–17777. - PubMed
-
- Blaser MJ. The bacteria behind ulcers. Sci Am. 1996;274:92–97. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
