

Polymerase chain reaction selects a novel disintegrin proteinase from CD40-activated germinal center dendritic cells
- PMID: 9271581
- PMCID: PMC2199019
- DOI: 10.1084/jem.186.5.655
Polymerase chain reaction selects a novel disintegrin proteinase from CD40-activated germinal center dendritic cells
Abstract
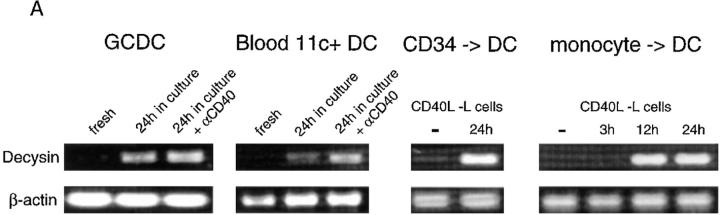
To identify genes expressed by a specific subset of dendritic cells found in vivo a polymerase chain reaction-based cDNA subtraction technique was applied to the recently described germinal center dendritic cells. A novel member of the disintegrin metalloproteinase family was cloned which comprises a not typical zinc-chelating catalytic site most similar to a bacterial metalloproteinase. Dendritic cell precursors or immature dendritic cells express no or low levels of the message. It is induced to high levels upon spontaneous or CD40-dependent maturation and in a mixed lymphocyte reaction. In situ hybridization showed distinct expression of this gene in the germinal center. This, together with the findings that certain disintegrin metalloproteinases regulate the activity of tumor necrosis factor alpha and that metalloproteinases have also been implicated in FasL processing, suggest that this novel molecule may play an important role in dendritic cell function and their interactions with germinal center T cells.
Figures








References
-
- Cella M, Sallusto F, Lanzavecchia A. Origin, maturation and antigen presenting function of dendritic cells. Curr Opin Immunol. 1997;9:10–16. - PubMed
-
- Caux, C., and J. Banchereau. 1996. In vitro regulation of dendritic cell development and function. In Blood Cell Biochemistry. T. Whetton and J. Gordon, editors. Plenum Press, London. 263–301.
-
- Grouard G, Durand I, Filgueira L, Banchereau J, Liu YJ. Dendritic cells capable of stimulating T cells in germinal centers. Nature (Lond) 1996;384:364–367. - PubMed
-
- Caux C, Dezutter-Dambuyant C, Schmitt D, Banchereau J. GM-CSF and TNF-α cooperate in the generation of dendritic Langerhans cells. Nature (Lond) 1992;360:258–261. - PubMed
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
