

High levels of a major histocompatibility complex II-self peptide complex on dendritic cells from the T cell areas of lymph nodes
- PMID: 9271582
- PMCID: PMC2199017
- DOI: 10.1084/jem.186.5.665
High levels of a major histocompatibility complex II-self peptide complex on dendritic cells from the T cell areas of lymph nodes
Abstract
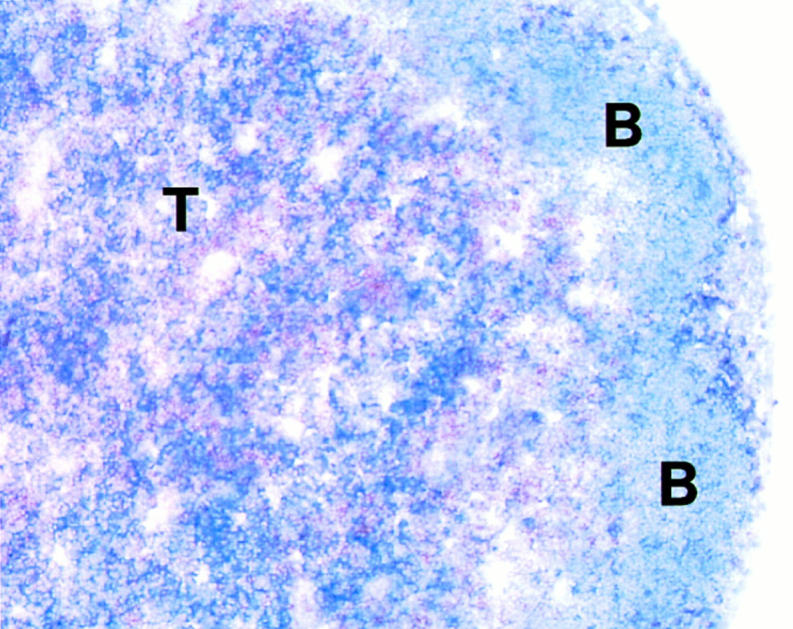
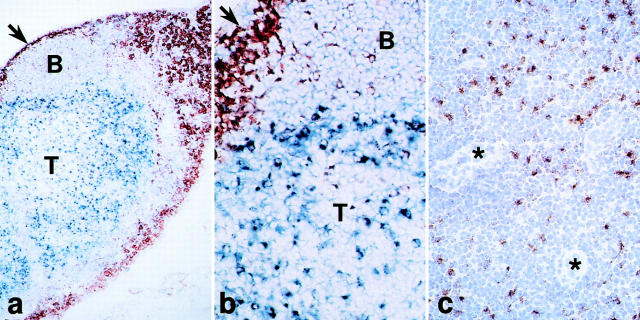
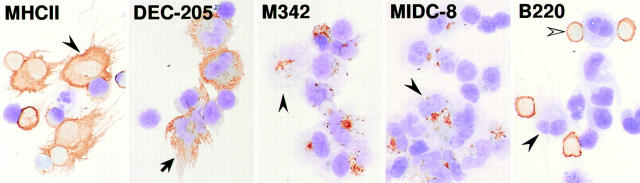
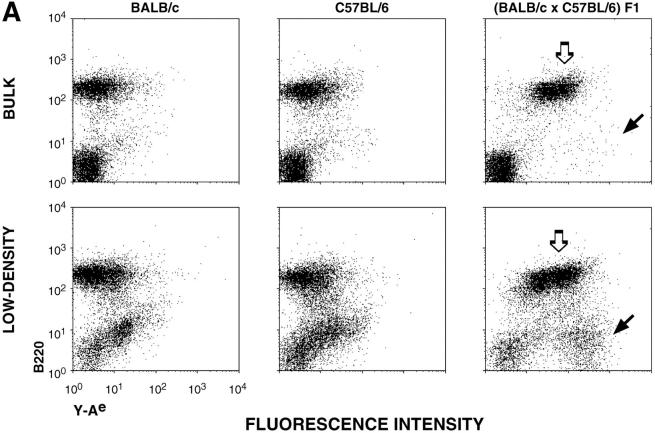
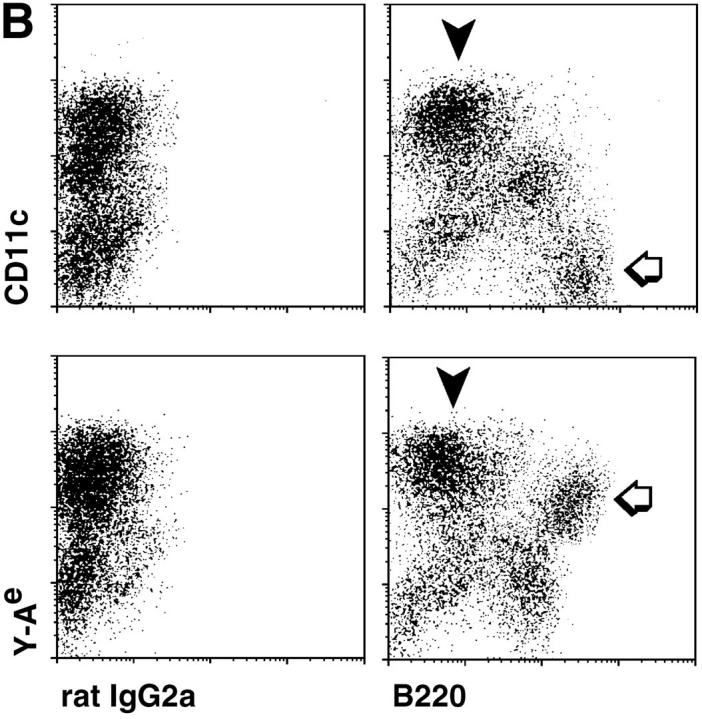
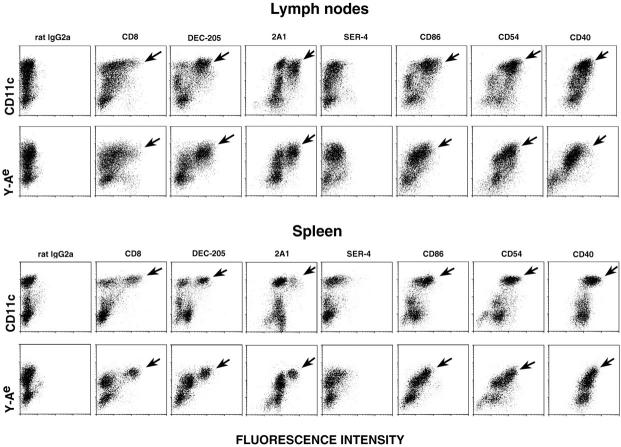
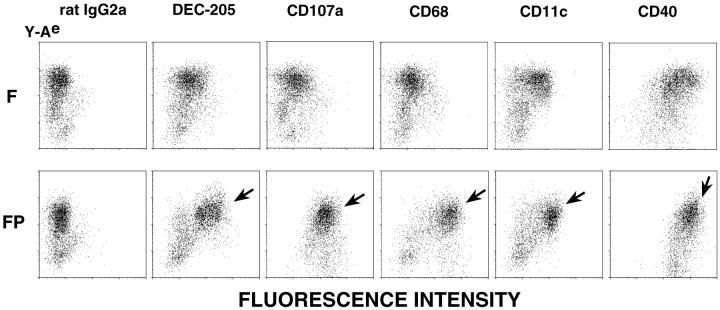
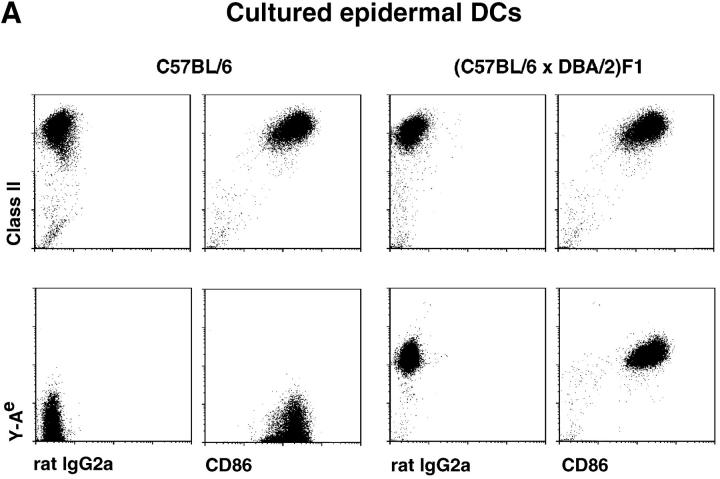
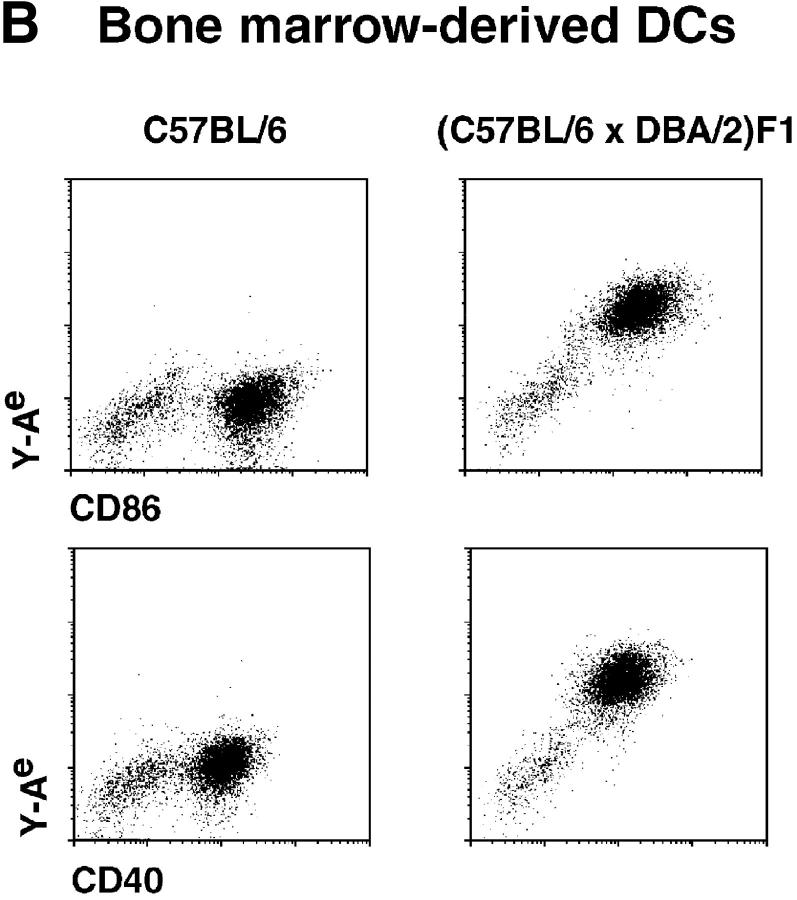
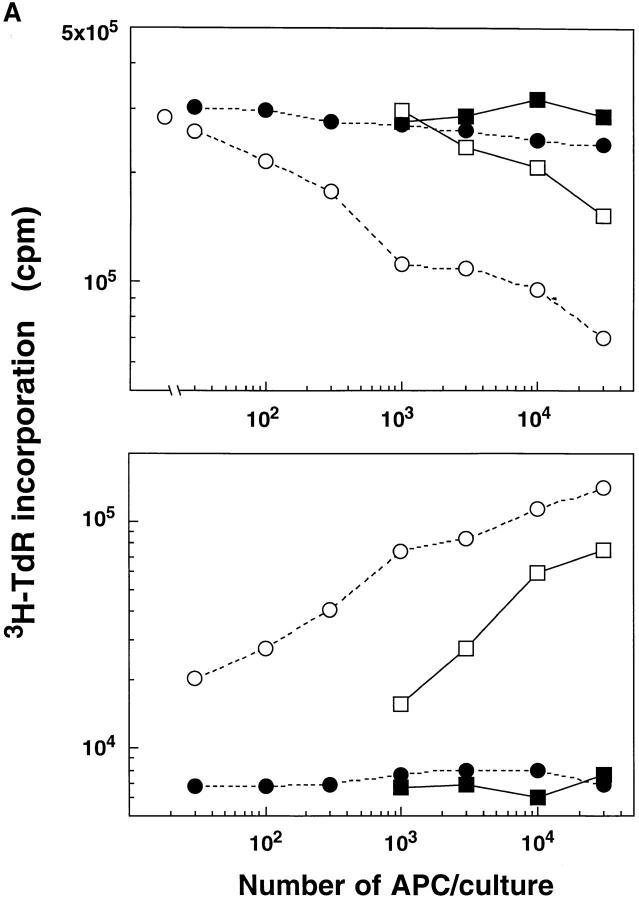
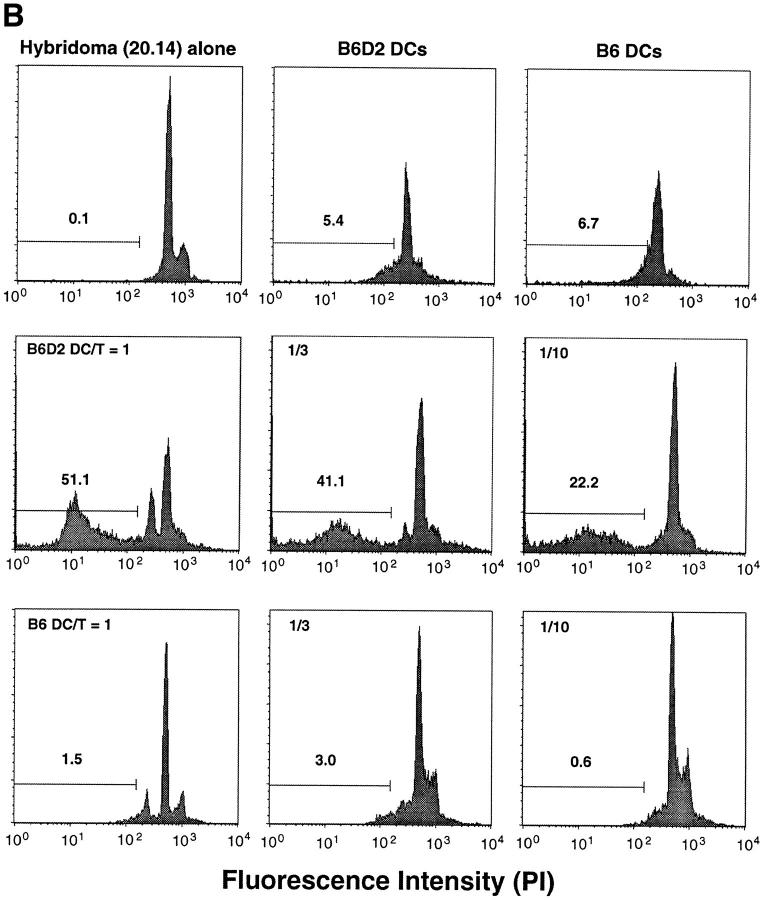
T lymphocytes recirculate continually through the T cell areas of peripheral lymph nodes. During each passage, the T cells survey the surface of large dendritic cells (DCs), also known as interdigitating cells. However, these DCs have been difficult to release from the lymph node. By emphasizing the use of calcium-free media, as shown by Vremec et al. (Vremec, D., M. Zorbas, R. Scollay, D.J. Saunders, C.F. Ardavin, L. Wu, and K. Shortman. 1992. J. Exp. Med. 176:47-58.), we have been able to release and enrich DCs from the T cell areas. The DCs express the CD11c leukocyte integrin, the DEC-205 multilectin receptor for antigen presentation, the intracellular granule antigens which are recognized by monoclonal antibodies M342, 2A1, and MIDC-8, very high levels of MHC I and MHC II, and abundant accessory molecules such as CD40, CD54, and CD86. When examined with the Y-Ae monoclonal which recognizes complexes formed between I-Ab and a peptide derived from I-Ealpha, the T cell area DCs expressed the highest levels. The enriched DCs also stimulated a T-T hybridoma specific for this MHC II-peptide complex, and the hybridoma underwent apoptosis. Therefore DCs within the T cell areas can be isolated. Because they present very high levels of self peptides, these DCs should be considered in the regulation of self reactivity in the periphery.
Figures
References
-
- Liu Y-J, Grouard G, de Bouteiller O, Banchereau J. Follicular dendritic cells and germinal centers. Int Rev Cytol. 1996;166:139–179. - PubMed
-
- Szakal AK, Kosco MH, Tew JG. Microanatomy of lymphoid tissue during humoral immune responses: structure function relationships. Annu Rev Immunol. 1989;7:91–109. - PubMed
-
- Steinman RM. The dendritic cell system and its role in immunogenicity. Annu Rev Immunol. 1991;9:271–296. - PubMed
-
- Williams LA, Egner W, Hart DN. Isolation and function of human dendritic cells. Int Rev Cytol. 1994;153:41–103. - PubMed
-
- Steinman RM, Pack M, Inaba K. Dendritic cells in the T cell areas of lymphoid organs. Immunol Rev. 1997;156:25–37. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
