

Spinophilin, a novel protein phosphatase 1 binding protein localized to dendritic spines
- PMID: 9275233
- PMCID: PMC23308
- DOI: 10.1073/pnas.94.18.9956
Spinophilin, a novel protein phosphatase 1 binding protein localized to dendritic spines
Abstract
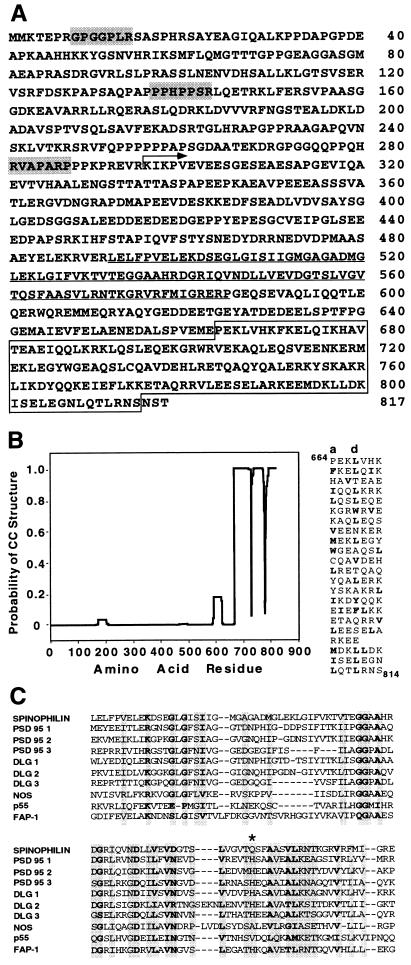
Dendritic spines receive the vast majority of excitatory synaptic contacts in the mammalian brain and are presumed to contain machinery for the integration of various signal transduction pathways. Protein phosphatase 1 (PP1) is greatly enriched in dendritic spines and has been implicated in both the regulation of ionic conductances and long-term synaptic plasticity. The molecular mechanism whereby PP1 is localized to spines is unknown. We have now characterized a novel protein that forms a complex with the catalytic subunit of PP1 and is a potent modulator of PP1 enzymatic activity in vitro. Within the brain this protein displays a remarkably distinct localization to the heads of dendritic spines and has therefore been named spinophilin. Spinophilin has the properties expected of a scaffolding protein localized to the cell membrane and contains a single consensus sequence in PSD95/DLG/zo-1, which implies cross-linking of PP1 to transmembrane protein complexes. We propose that spinophilin represents a novel targeting subunit for PP1, which directs the enzyme to those substrates in the dendritic spine compartment, e.g., neurotransmitter receptors, which mediate the regulation of synaptic function by PP1.
Figures
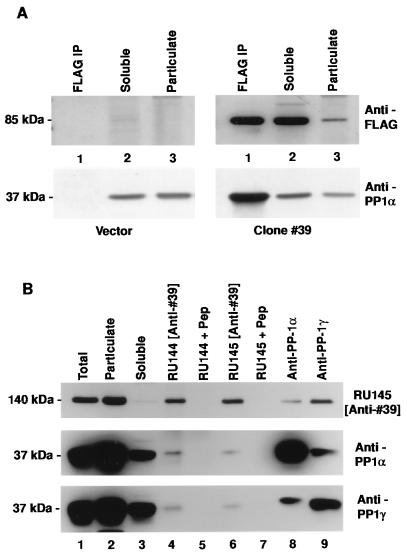
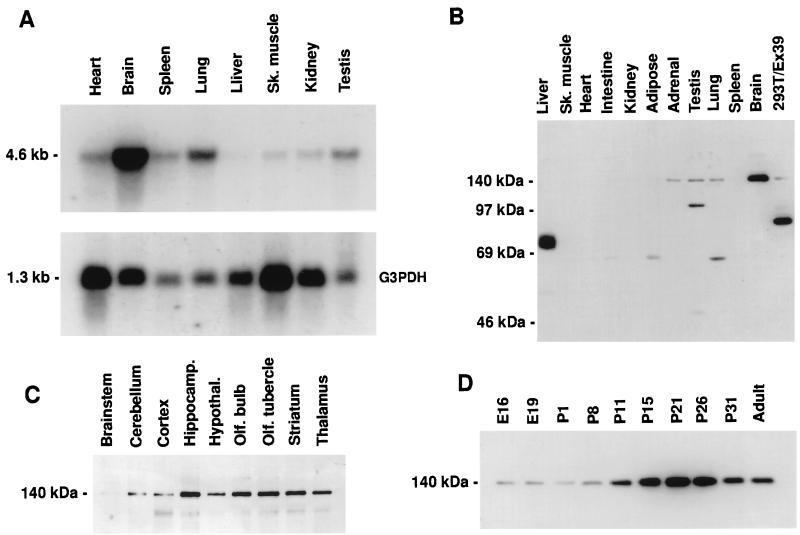
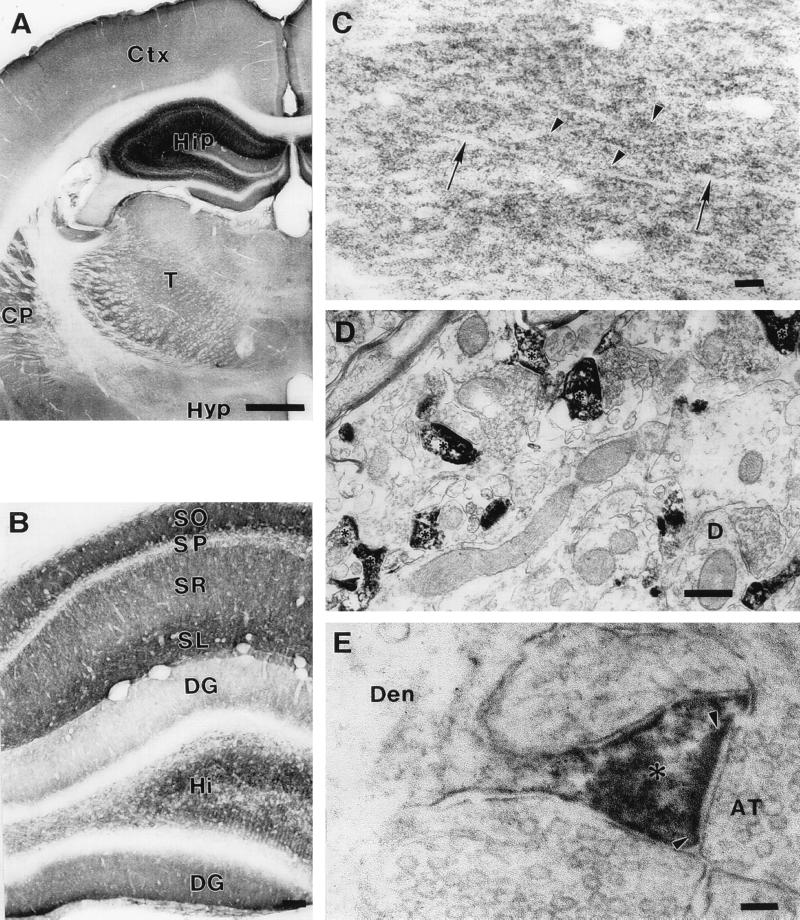
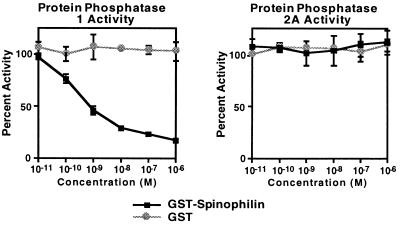
References
-
- Harris K M, Kater S B. Annu Rev Neurosci. 1994;17:341–371. - PubMed
-
- Surmeier D J, Bargas J, Hemmings H C, Jr, Nairn A C, Greengard P. Neuron. 1995;14:385–397. - PubMed
-
- Endo S, Critz S D, Byrne J H, Shenolikar S. J Neurochem. 1995;64:1833–1840. - PubMed
-
- Mulkey R M, Endo S, Shenolikar S, Malenka R C. Nature (London) 1994;369:486–488. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
