

Adrenaline stimulates glucagon secretion in pancreatic A-cells by increasing the Ca2+ current and the number of granules close to the L-type Ca2+ channels
- PMID: 9276750
- PMCID: PMC2229364
- DOI: 10.1085/jgp.110.3.217
Adrenaline stimulates glucagon secretion in pancreatic A-cells by increasing the Ca2+ current and the number of granules close to the L-type Ca2+ channels
Abstract
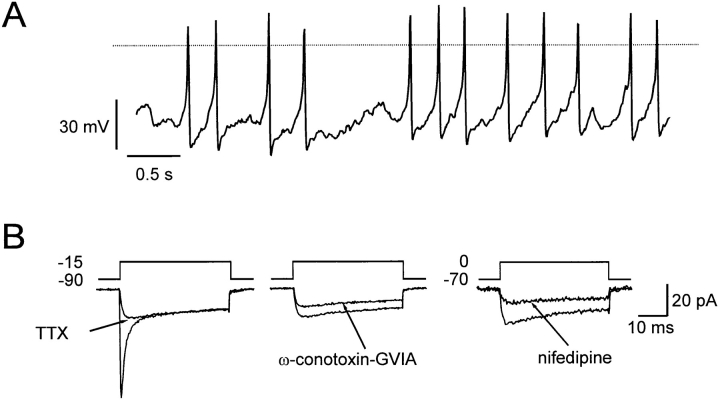
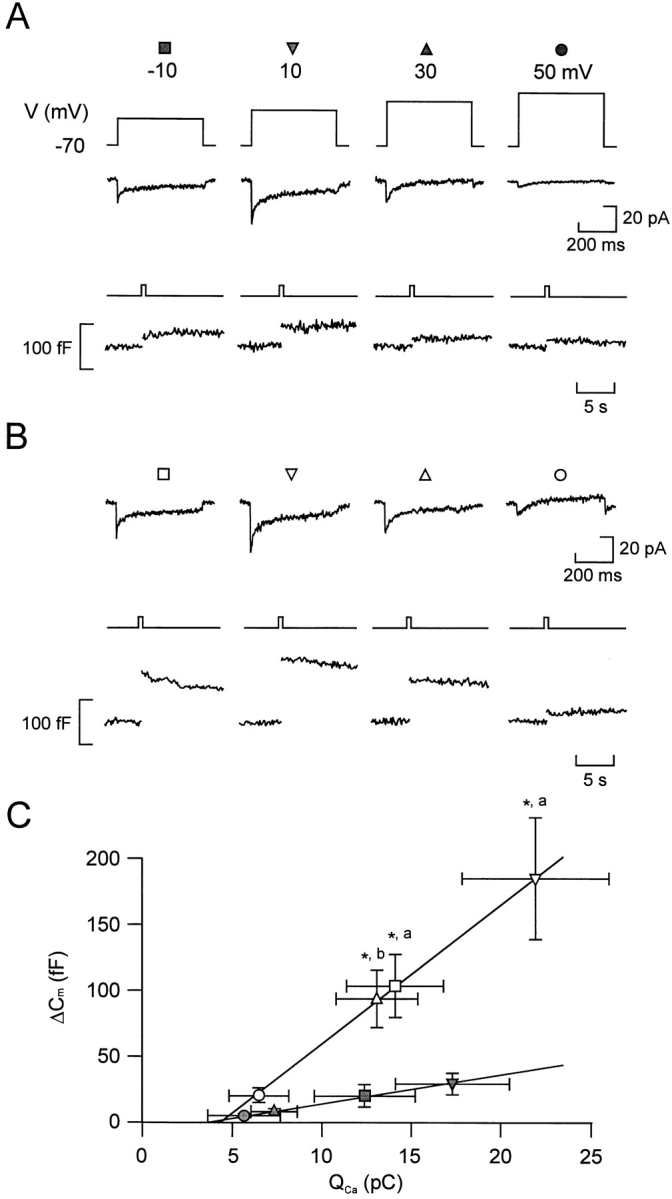
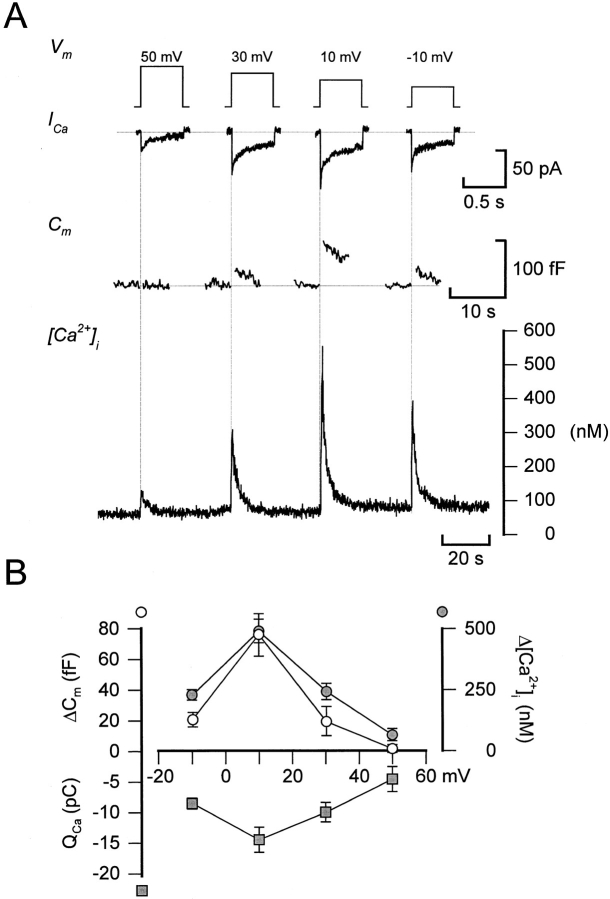
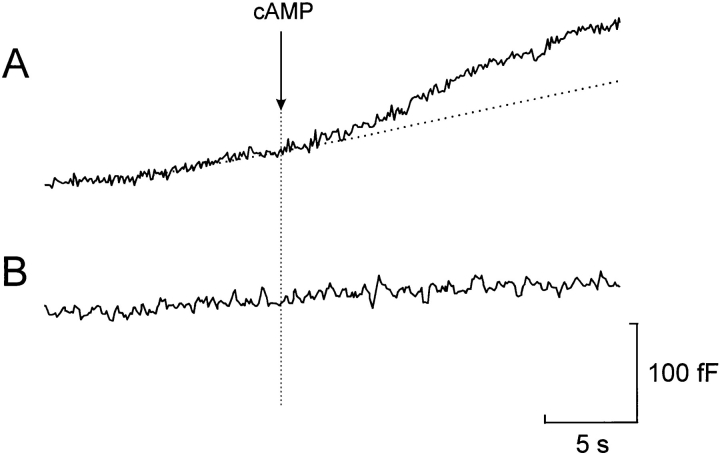
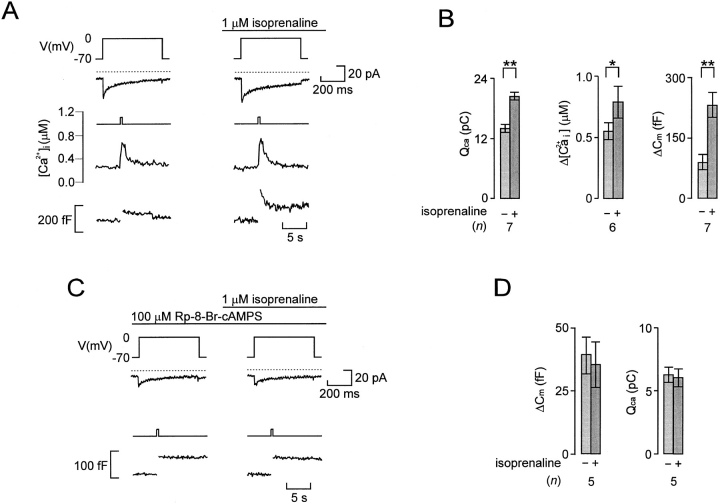
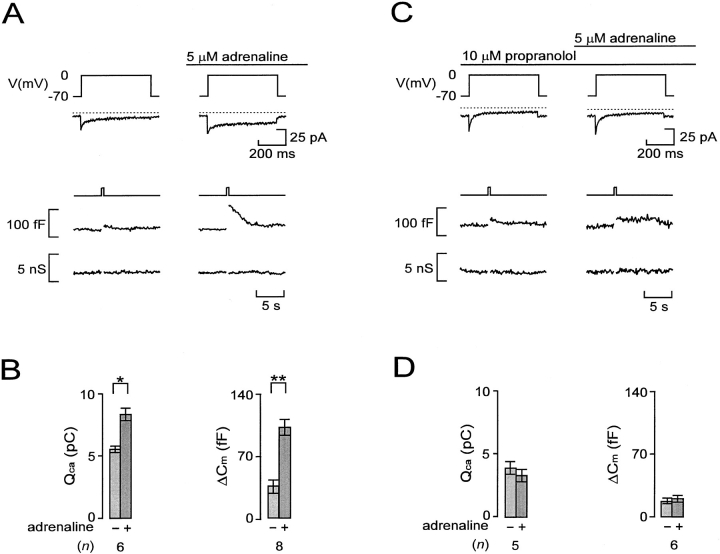
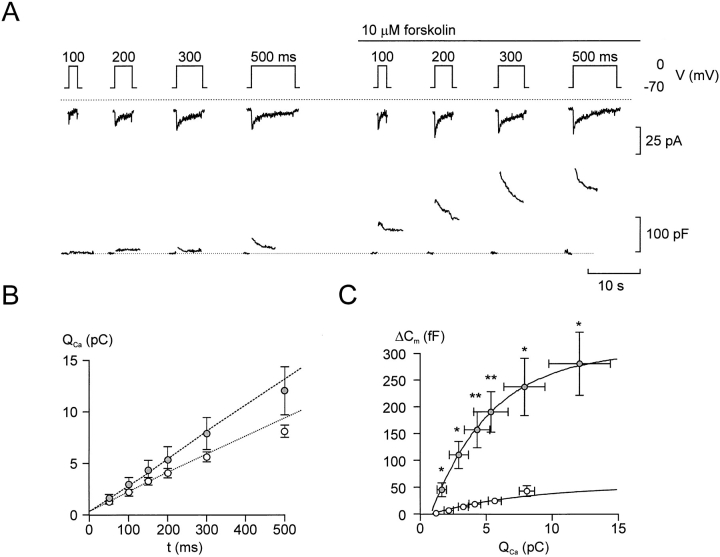
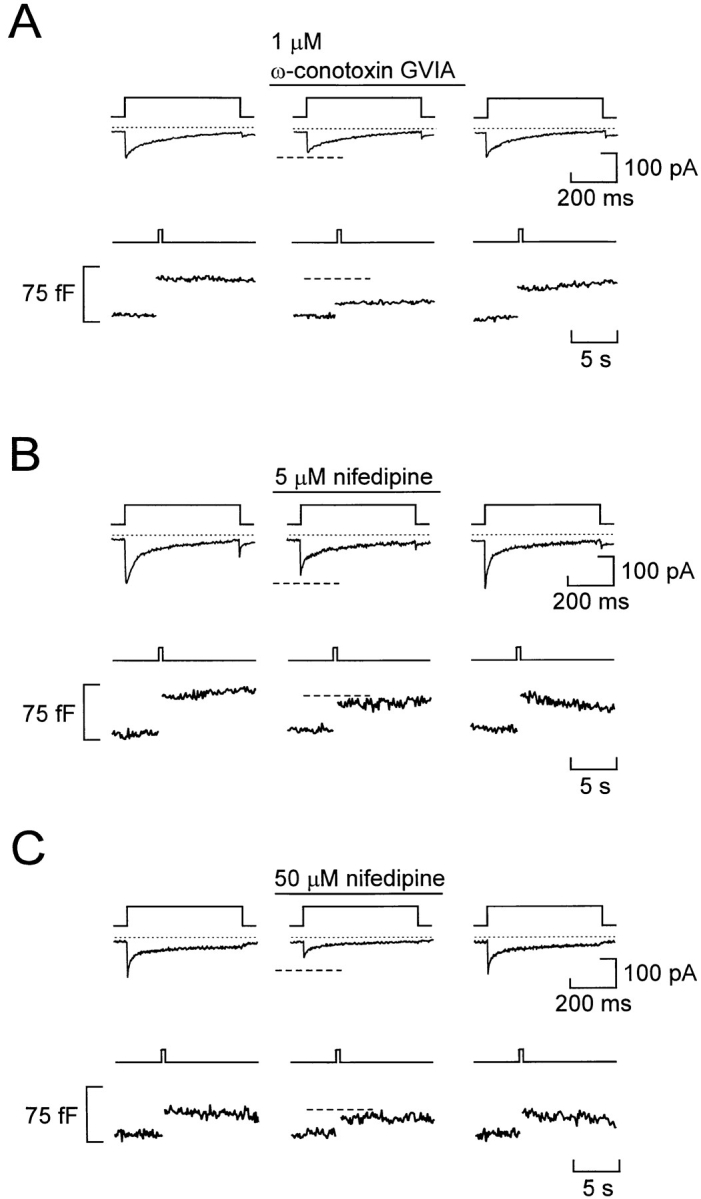
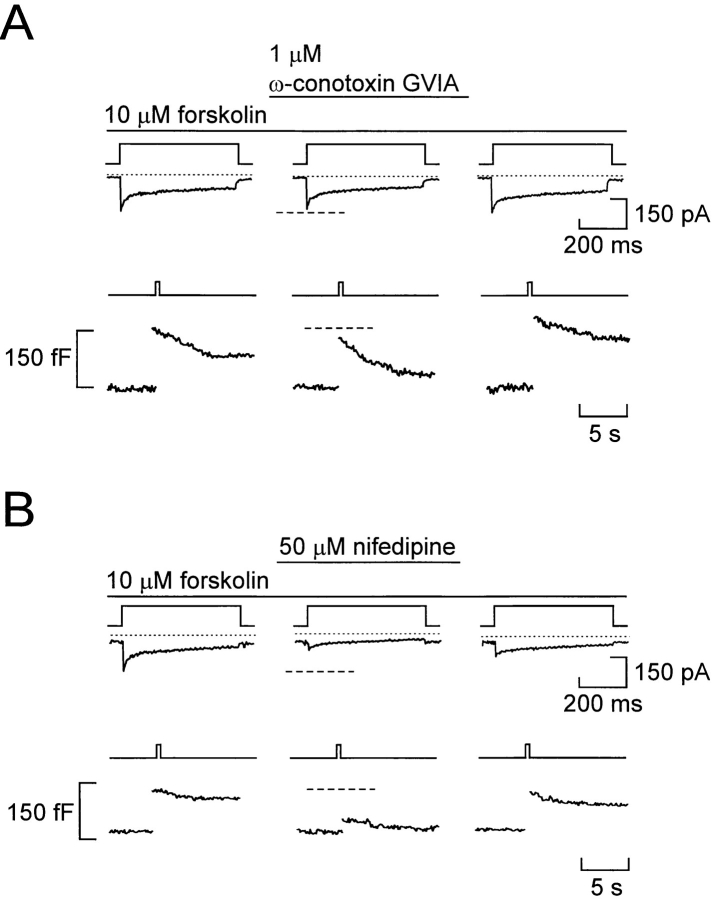
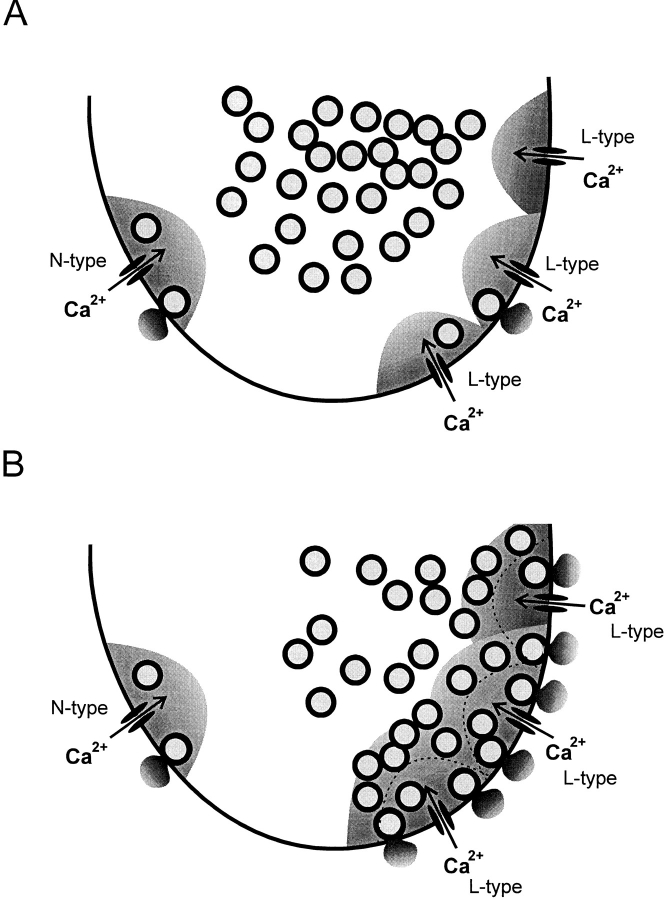
We have monitored electrical activity, voltage-gated Ca2+ currents, and exocytosis in single rat glucagon-secreting pancreatic A-cells. The A-cells were electrically excitable and generated spontaneous Na+- and Ca2+-dependent action potentials. Under basal conditions, exocytosis was tightly linked to Ca2+ influx through omega-conotoxin-GVIA-sensitive (N-type) Ca2+ channels. Stimulation of the A-cells with adrenaline (via beta-adrenergic receptors) or forskolin produced a greater than fourfold PKA-dependent potentiation of depolarization-evoked exocytosis. This enhancement of exocytosis was due to a 50% enhancement of Ca2+ influx through L-type Ca2+ channels, an effect that accounted for <30% of the total stimulatory action. The remaining 70% of the stimulation was attributable to an acceleration of granule mobilization resulting in a fivefold increase in the number of readily releasable granules near the L-type Ca2+ channels.
Figures
References
-
- Ahrén B, Stern JS, Gingerich RL, Curry DL, Havel PJ. Glucagon secretory response to hypoglycemia, adrenaline, and carbachol in streptozotocin-diabetic rats. Acta Physiol Scand. 1995;155:215–221. - PubMed
-
- Ämmälä C, Ashcroft FM, Rorsman P. Calcium-independent potentiation of insulin release by cyclic AMP in single β-cells. Nature (Lond) 1993a;363:356–358. - PubMed
-
- Artalejo CR, Adams ME, Fox AP. Three types of Ca2+channel trigger secretion with different efficacies in chromaffin cells. Nature (Lond) 1994;367:72–76. - PubMed
-
- Berts A, Gylfe E, Hellman B. Ca2+oscillations in pancreatic islet cells secreting glucagon and somatostatin. Biochem Biophys Res Commun. 1995;208:644–649. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
