

Mitotic spindle poles are organized by structural and motor proteins in addition to centrosomes
- PMID: 9281583
- PMCID: PMC2136753
- DOI: 10.1083/jcb.138.5.1055
Mitotic spindle poles are organized by structural and motor proteins in addition to centrosomes
Abstract
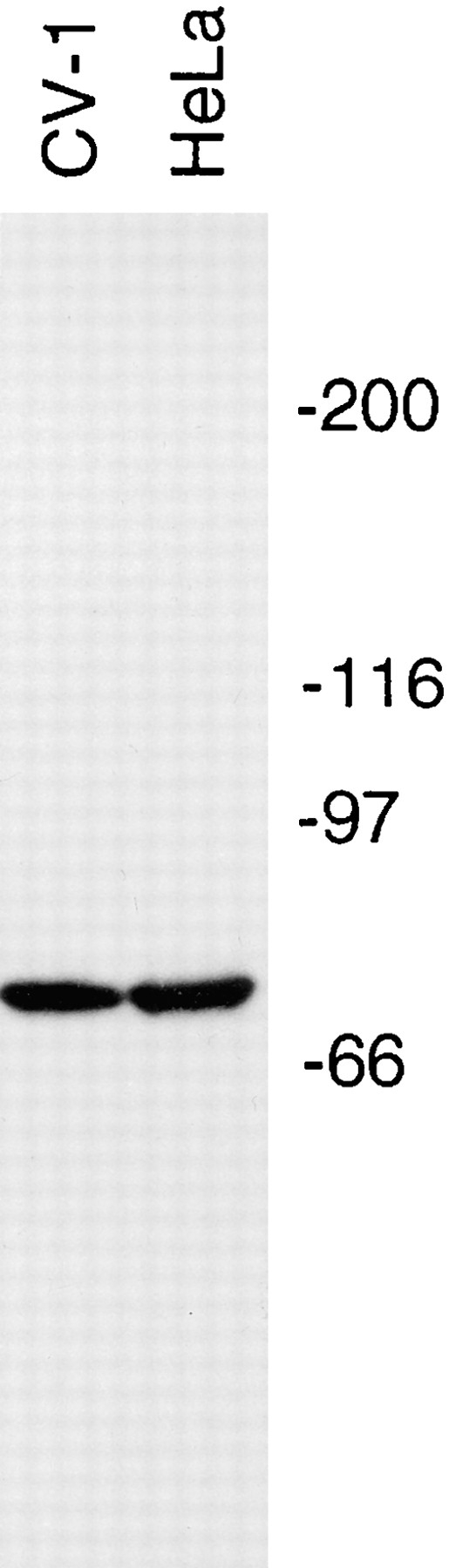
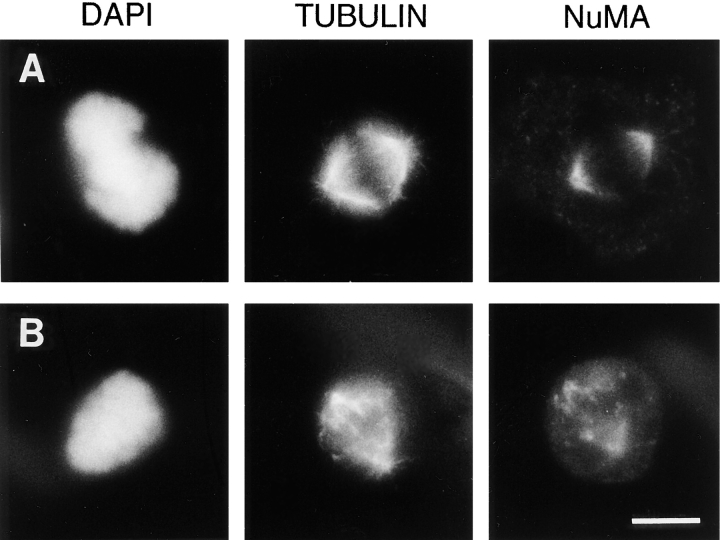
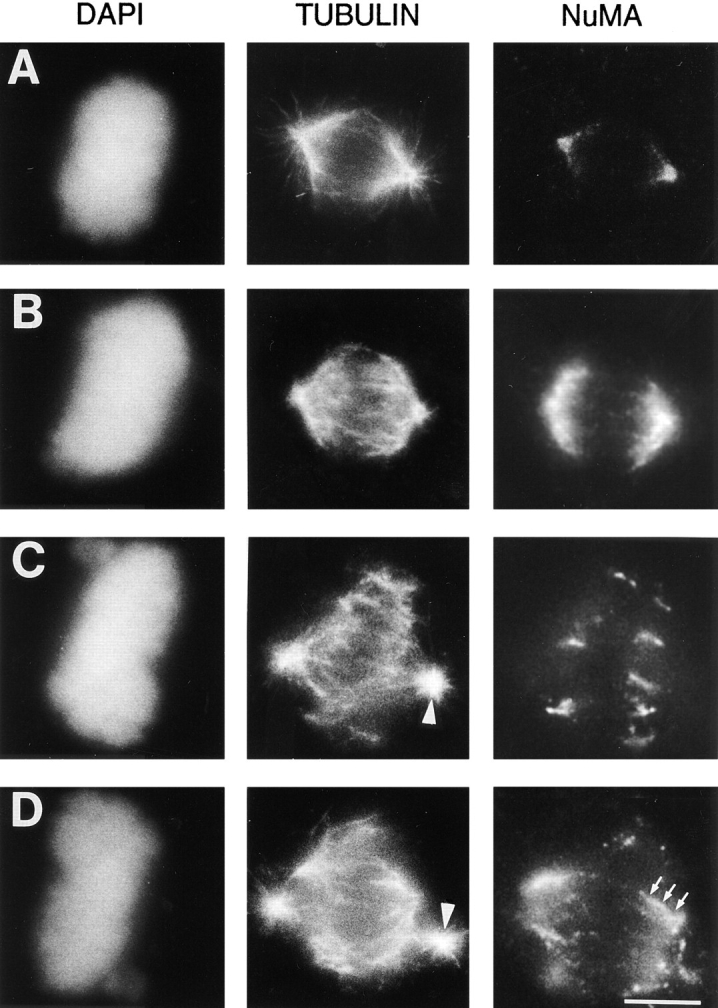
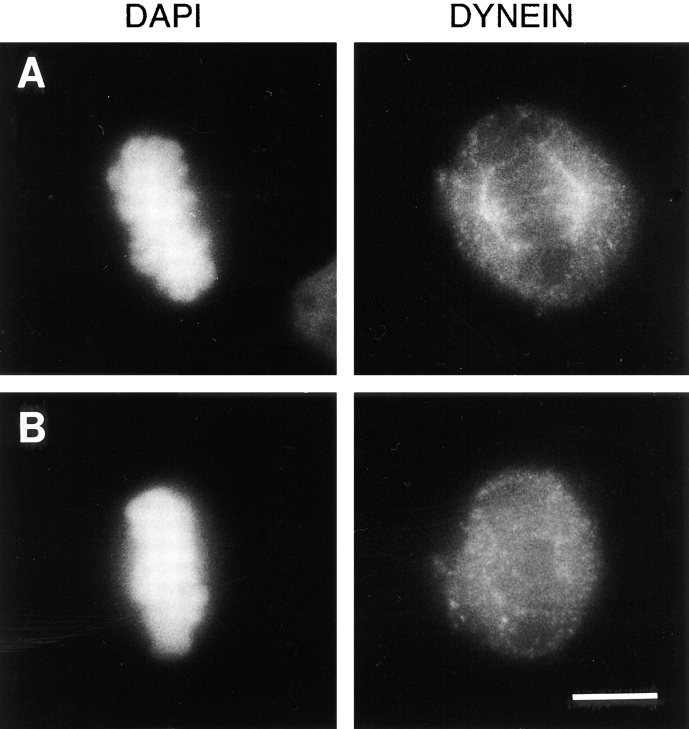
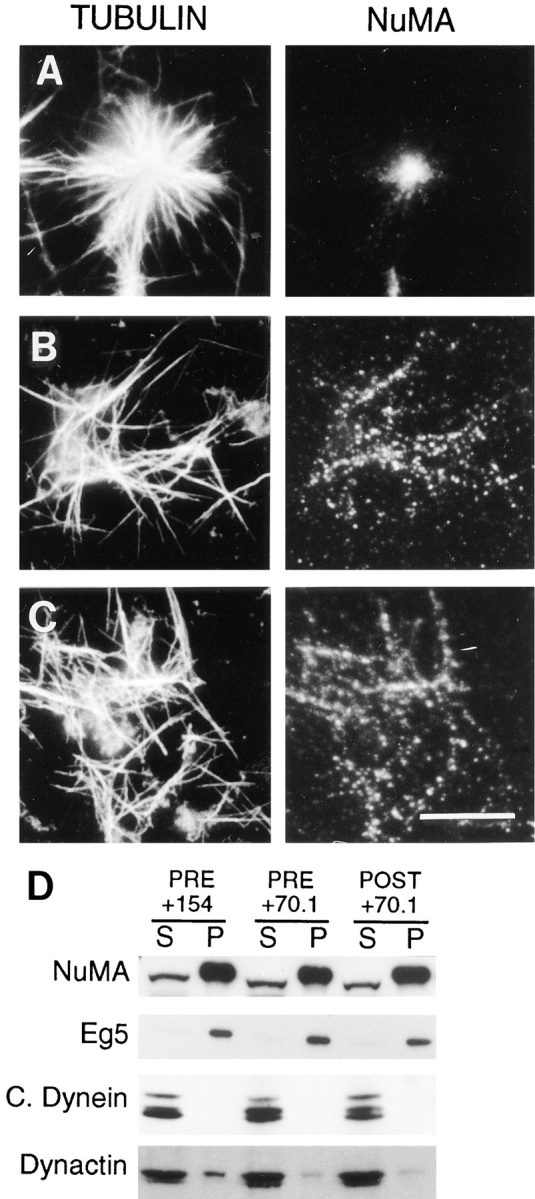
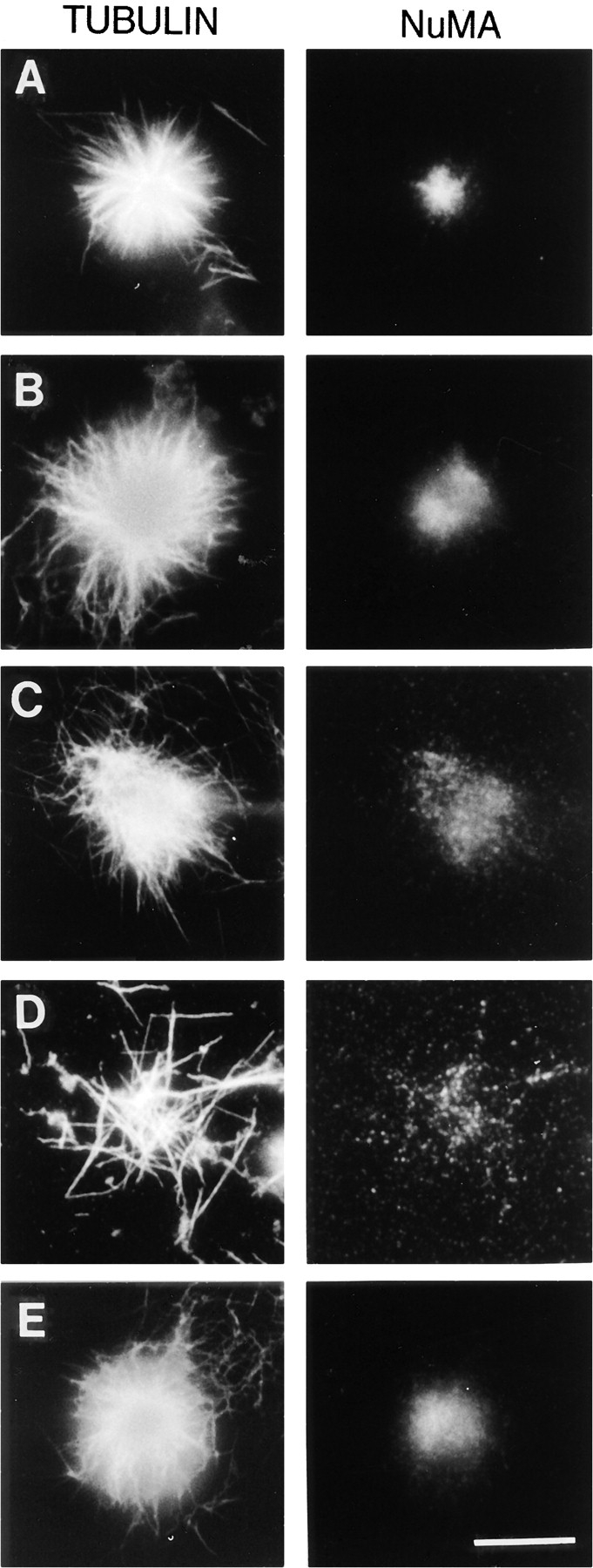
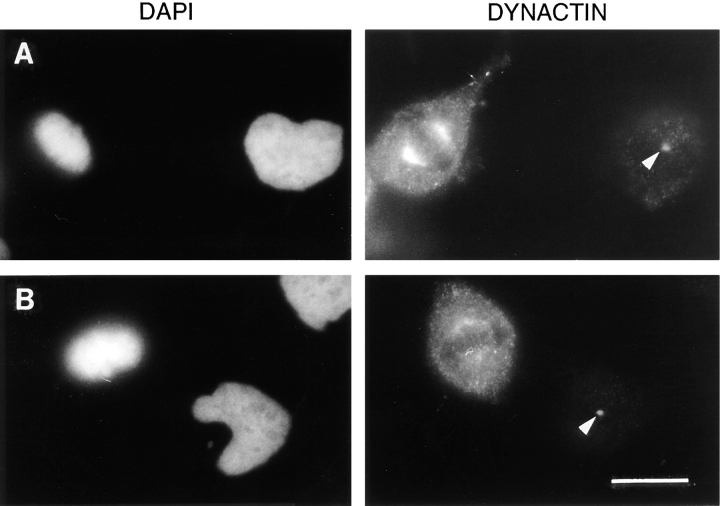
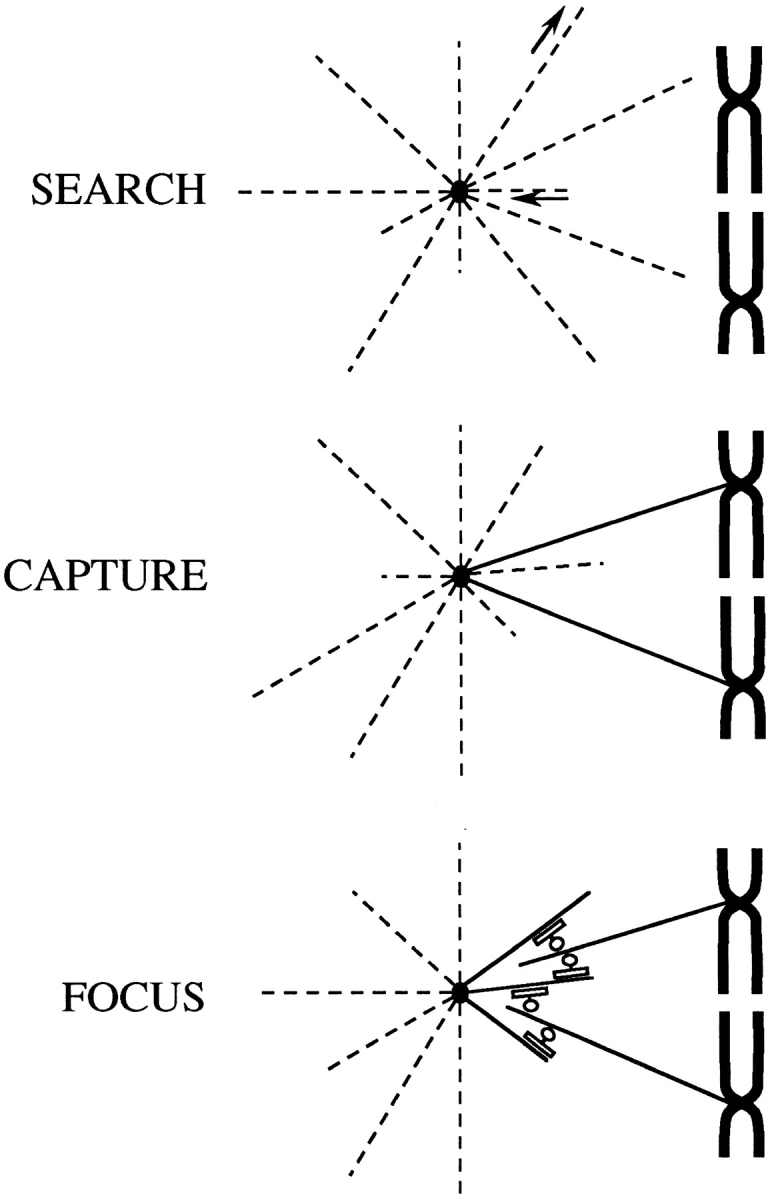
The focusing of microtubules into mitotic spindle poles in vertebrate somatic cells has been assumed to be the consequence of their nucleation from centrosomes. Contrary to this simple view, in this article we show that an antibody recognizing the light intermediate chain of cytoplasmic dynein (70.1) disrupts both the focused organization of microtubule minus ends and the localization of the nuclear mitotic apparatus protein at spindle poles when injected into cultured cells during metaphase, despite the presence of centrosomes. Examination of the effects of this dynein-specific antibody both in vitro using a cell-free system for mitotic aster assembly and in vivo after injection into cultured cells reveals that in addition to its direct effect on cytoplasmic dynein this antibody reduces the efficiency with which dynactin associates with microtubules, indicating that the antibody perturbs the cooperative binding of dynein and dynactin to microtubules during spindle/aster assembly. These results indicate that microtubule minus ends are focused into spindle poles in vertebrate somatic cells through a mechanism that involves contributions from both centrosomes and structural and microtubule motor proteins. Furthermore, these findings, together with the recent observation that cytoplasmic dynein is required for the formation and maintenance of acentrosomal spindle poles in extracts prepared from Xenopus eggs (Heald, R., R. Tournebize, T. Blank, R. Sandaltzopoulos, P. Becker, A. Hyman, and E. Karsenti. 1996. Nature (Lond.). 382: 420-425) demonstrate that there is a common mechanism for focusing free microtubule minus ends in both centrosomal and acentrosomal spindles. We discuss these observations in the context of a search-capture-focus model for spindle assembly.
Figures
References
-
- Allan V. Motor proteins: a dynamic duo. Curr Biol. 1996;6:630–633. - PubMed
-
- Bastmeyer M, Steffen W, Fuge H. Immunostaining of spindle components in tipulid spermatocytes using a serum against pericentriolar material. Eur J Cell Biol. 1986;42:305–310. - PubMed
-
- Belmont LD, Hyman AA, Sawin KE, Mitchison TJ. Real-time visualization of cell cycle-dependent changes in microtubule dynamics in cytoplasmic extracts. Cell. 1990;62:579–589. - PubMed
-
- Blangy A, Lane HA, d'Herin P, Harper M, Kress M, Nigg EA. Phosphorylation by p34cdc2 regulates spindle association of human Eg5, a kinesin-related motor essential for bipolar spindle formation in vivo. Cell. 1995;83:1159–1169. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
