

Rapid effector function in CD8+ memory T cells
- PMID: 9294140
- PMCID: PMC2199056
- DOI: 10.1084/jem.186.6.859
Rapid effector function in CD8+ memory T cells
Abstract
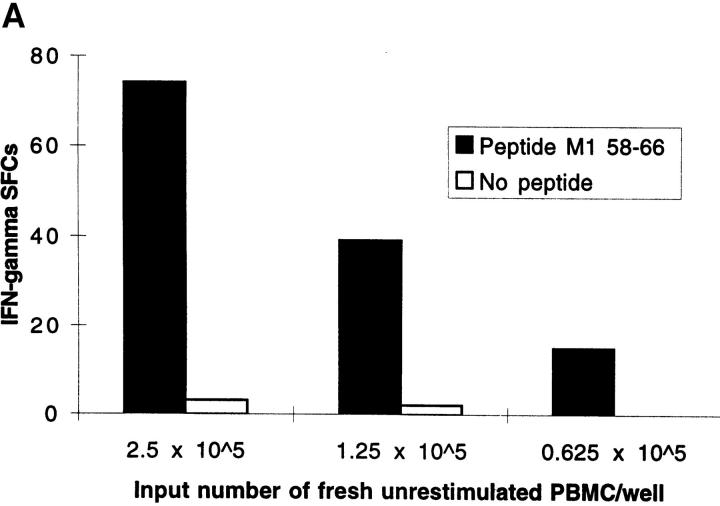
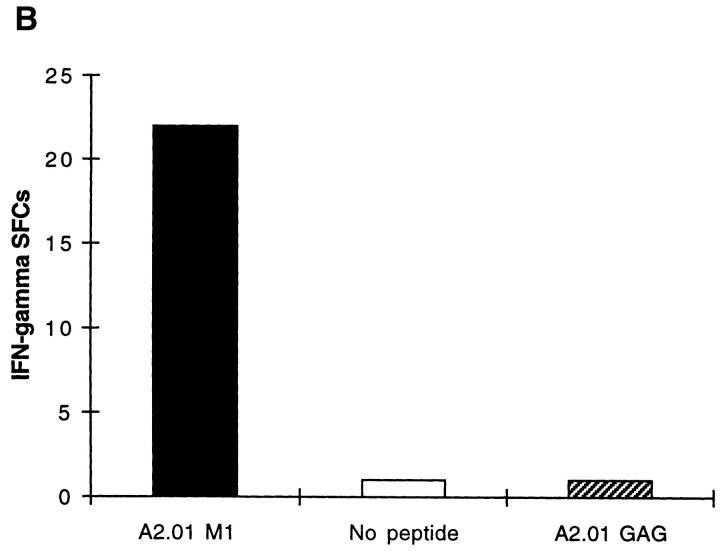
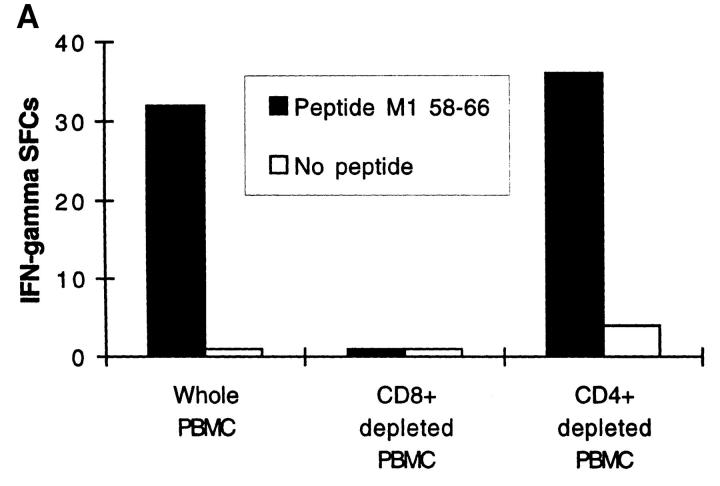
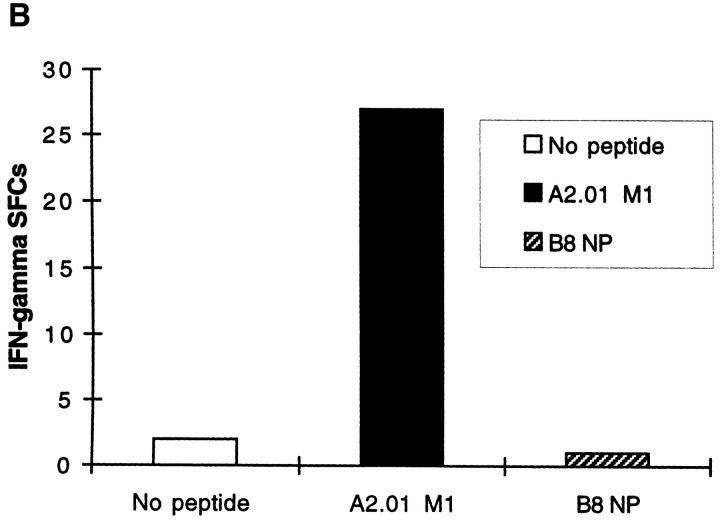
The nature of the CD8+ T cells that underlie antiviral protective immunological memory in vivo is unclear. We have characterized peptide-specific CD8+ T lymphocytes directly ex vivo from peripheral blood in humans with past exposure to influenza virus, using single cell interferon gamma (IFN-gamma) release as a measure of effector function. In individuals in the memory state with respect to influenza virus infection, unrestimulated antigen-specific CD8+ T cells displayed IFN-gamma release within 6 h of antigen contact, identifying a population of memory CD8+ T cells that exhibit effector function without needing to divide and differentiate over several days. We have quantified circulating CD8+ effector T cells specific for six different MHC class I-restricted influenza virus epitopes. Enumeration of these CD8+ T cells gives frequencies of peptide-specific T cells that correlate with, but are in general severalfold higher than, CTL precursor frequencies derived from limiting dilution analysis, indicating that this novel population of memory CD8+ T cells has hitherto been undetected by standard means. The phenotype of these cells, which persist at a low frequency long after recovery from an acute viral infection, suggests that they play a role in protective immunological memory.
Figures
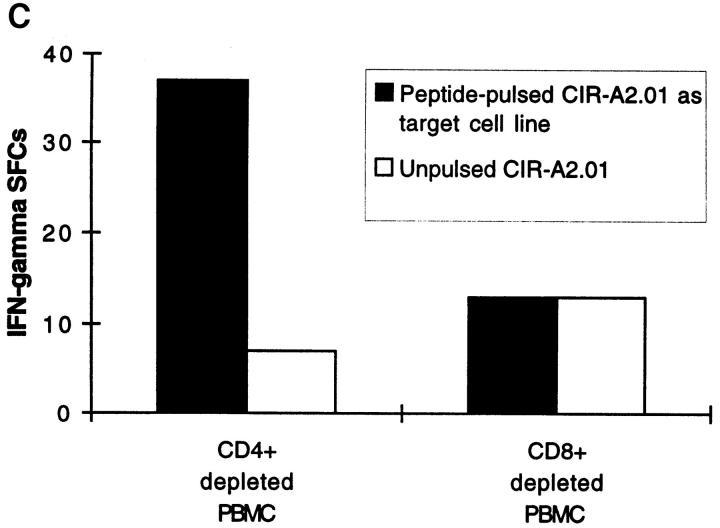
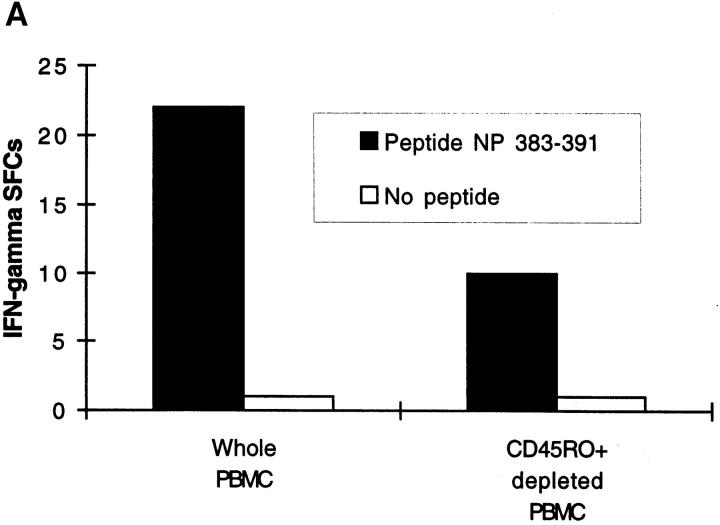
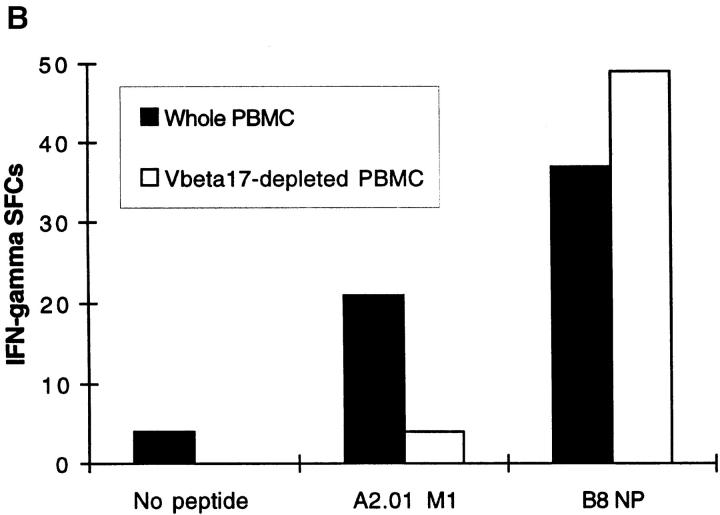
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
