

Control of time-dependent biological processes by temporally patterned input
- PMID: 9294230
- PMCID: PMC23382
- DOI: 10.1073/pnas.94.19.10444
Control of time-dependent biological processes by temporally patterned input
Abstract
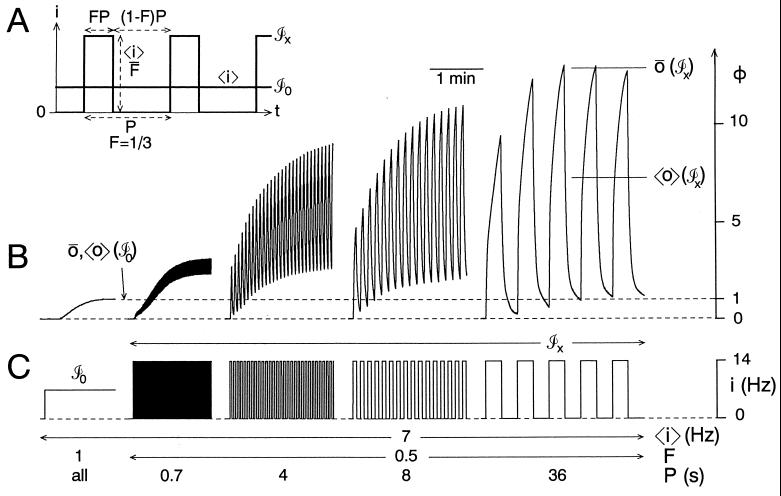
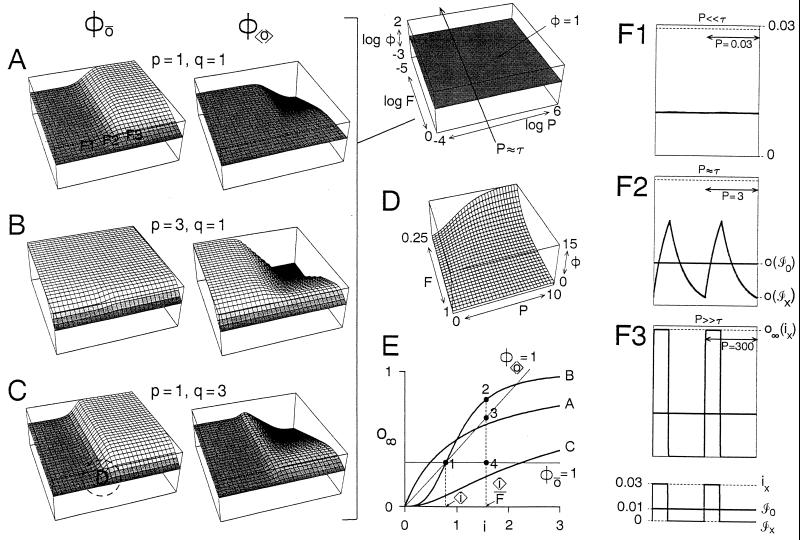
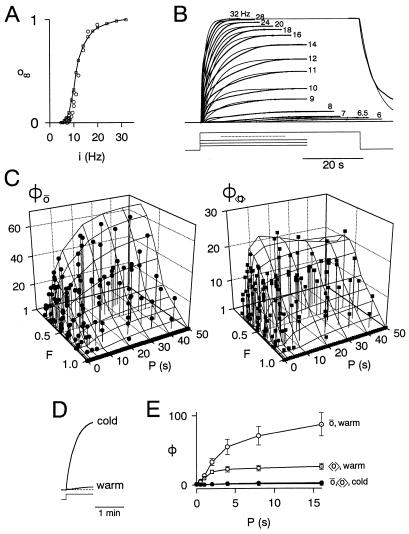
Temporal patterning of biological variables, in the form of oscillations and rhythms on many time scales, is ubiquitous. Altering the temporal pattern of an input variable greatly affects the output of many biological processes. We develop here a conceptual framework for a quantitative understanding of such pattern dependence, focusing particularly on nonlinear, saturable, time-dependent processes that abound in biophysics, biochemistry, and physiology. We show theoretically that pattern dependence is governed by the nonlinearity of the input-output transformation as well as its time constant. As a result, only patterns on certain time scales permit the expression of pattern dependence, and processes with different time constants can respond preferentially to different patterns. This has implications for temporal coding and decoding, and allows differential control of processes through pattern. We show how pattern dependence can be quantitatively predicted using only information from steady, unpatterned input. To apply our ideas, we analyze, in an experimental example, how muscle contraction depends on the pattern of motorneuron firing.
Figures

Similar articles
-
Rate coding in a chain of pulse-coupled oscillators.Phys Rev E Stat Phys Plasmas Fluids Relat Interdiscip Topics. 1999 Oct;60(4 Pt B):4564-70. doi: 10.1103/physreve.60.4564. Phys Rev E Stat Phys Plasmas Fluids Relat Interdiscip Topics. 1999. PMID: 11970314
-
The neuromuscular transform: the dynamic, nonlinear link between motor neuron firing patterns and muscle contraction in rhythmic behaviors.J Neurophysiol. 2000 Jan;83(1):207-31. doi: 10.1152/jn.2000.83.1.207. J Neurophysiol. 2000. PMID: 10634868
-
Temperature dependence and temperature compensation of kinetics of chemical oscillations; Belousov-Zhabotinskii reaction, glycolysis and circadian rhythms.J Theor Biol. 2008 Jan 7;250(1):103-12. doi: 10.1016/j.jtbi.2007.08.029. Epub 2007 Sep 6. J Theor Biol. 2008. PMID: 17920634
-
Circadian clocks - the fall and rise of physiology.Nat Rev Mol Cell Biol. 2005 Dec;6(12):965-71. doi: 10.1038/nrm1766. Nat Rev Mol Cell Biol. 2005. PMID: 16341082 Review.
-
Integration of biological clocks and rhythms.Compr Physiol. 2012 Apr;2(2):1213-39. doi: 10.1002/cphy.c100088. Compr Physiol. 2012. PMID: 23798299 Review.
Cited by
-
Muscle response to changing neuronal input in the lobster (Panulirus interruptus) stomatogastric system: slow muscle properties can transform rhythmic input into tonic output.J Neurosci. 1998 May 1;18(9):3433-42. doi: 10.1523/JNEUROSCI.18-09-03433.1998. J Neurosci. 1998. PMID: 9547250 Free PMC article.
-
Muscle anatomy is a primary determinant of muscle relaxation dynamics in the lobster (Panulirus interruptus) stomatogastric system.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2007 Nov;193(11):1101-13. doi: 10.1007/s00359-007-0261-7. Epub 2007 Aug 21. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2007. PMID: 17710408
-
Temporal pattern dependence of neuronal peptide transmitter release: models and experiments.J Neurosci. 2000 Sep 15;20(18):6760-72. doi: 10.1523/JNEUROSCI.20-18-06760.2000. J Neurosci. 2000. PMID: 10995819 Free PMC article.
-
Evaluation of gene association methods for coexpression network construction and biological knowledge discovery.PLoS One. 2012;7(11):e50411. doi: 10.1371/journal.pone.0050411. Epub 2012 Nov 30. PLoS One. 2012. PMID: 23226279 Free PMC article.
-
Variability of motor neuron spike timing maintains and shapes contractions of the accessory radula closer muscle of Aplysia.J Neurosci. 2006 Jun 28;26(26):7056-70. doi: 10.1523/JNEUROSCI.5277-05.2006. J Neurosci. 2006. PMID: 16807335 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
