

Rapid seizure-induced reduction of benzodiazepine and Zn2+ sensitivity of hippocampal dentate granule cell GABAA receptors
- PMID: 9295398
- PMCID: PMC2892718
- DOI: 10.1523/JNEUROSCI.17-19-07532.1997
Rapid seizure-induced reduction of benzodiazepine and Zn2+ sensitivity of hippocampal dentate granule cell GABAA receptors
Abstract
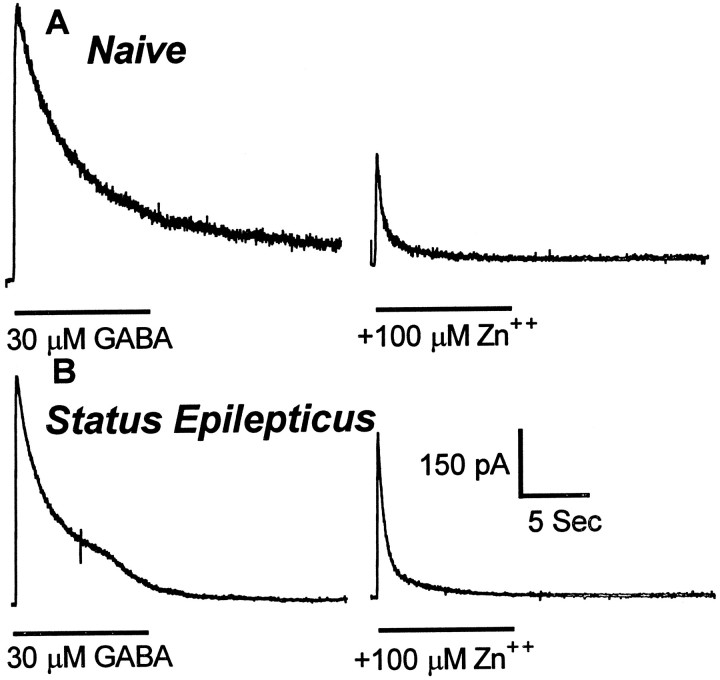
Fast synaptic inhibition in the forebrain is mediated primarily by GABA acting on GABAA receptors (GABARs). GABARs are regulated by numerous positive (barbiturates, benzodiazepines, and neurosteroids) and negative (picrotoxin, bicuculline, and Zn2+) allosteric modulators. The sensitivity of GABARs to GABA and to allosteric modulators changes gradually during normal development, during development of chronic epilepsy, and after prolonged exposure to GABAR agonists. Here we report the development of rapid functional plasticity of GABARs occurring over 45 min of continuous seizures (status epilepticus) in rats. Seizures induced in rats by administration of lithium followed by pilocarpine were readily terminated by the benzodiazepine diazepam when administered early during the seizures (after 10 min of seizures). However, during status epilepticus, there was a substantial reduction of diazepam potency for termination of the seizures. To determine whether the loss of sensitivity of the animals to diazepam was caused by an alteration of GABAR functional properties, we obtained whole-cell GABAR currents from hippocampal dentate granule cells isolated acutely from control rats and from rats undergoing status epilepticus. GABAR properties were characterized by determining GABA sensitivity and the sensitivity of GABARs to regulation by benzodiazepines, barbiturates, and Zn2+. When compared with those from naive controls, GABAR currents from rats undergoing status epilepticus were less sensitive to diazepam and Zn2+ but retained their sensitivity to GABA and pentobarbital. We conclude that the prolonged seizures of status epilepticus rapidly altered the functional properties of hippocampal dentate granule cell GABARs.
Figures









References
-
- Bertram EH, Lothman EW. Morphometric effects of intermittent kindled seizures and limbic status epilepticus in the dentate gyrus of the rat. Brain Res. 1993;603:25–31. - PubMed
-
- Buhl EH, Otis TS, Mody I. Zinc-induced collapse of augmented inhibition by GABA in a temporal lobe epilepsy model. Science. 1996;271:369–373. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical