

SOI1 encodes a novel, conserved protein that promotes TGN-endosomal cycling of Kex2p and other membrane proteins by modulating the function of two TGN localization signals
- PMID: 9314526
- PMCID: PMC2139830
- DOI: 10.1083/jcb.139.1.23
SOI1 encodes a novel, conserved protein that promotes TGN-endosomal cycling of Kex2p and other membrane proteins by modulating the function of two TGN localization signals
Abstract
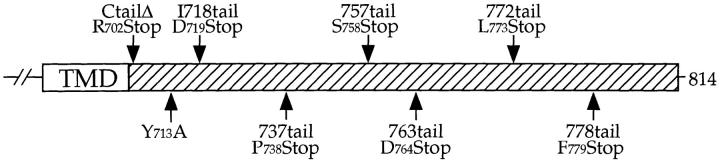
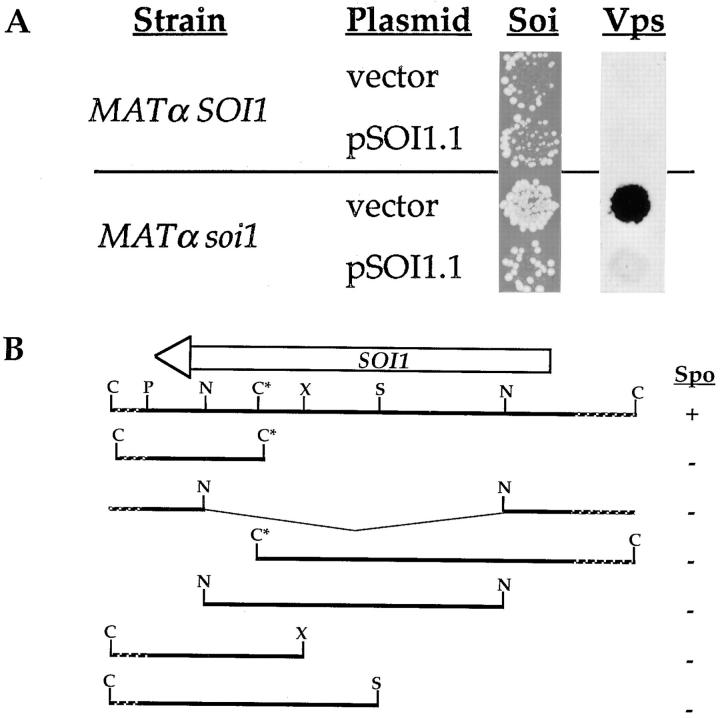
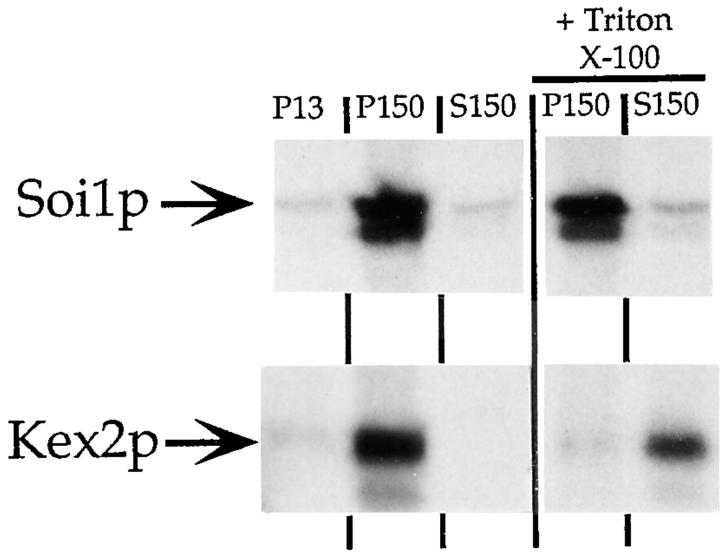
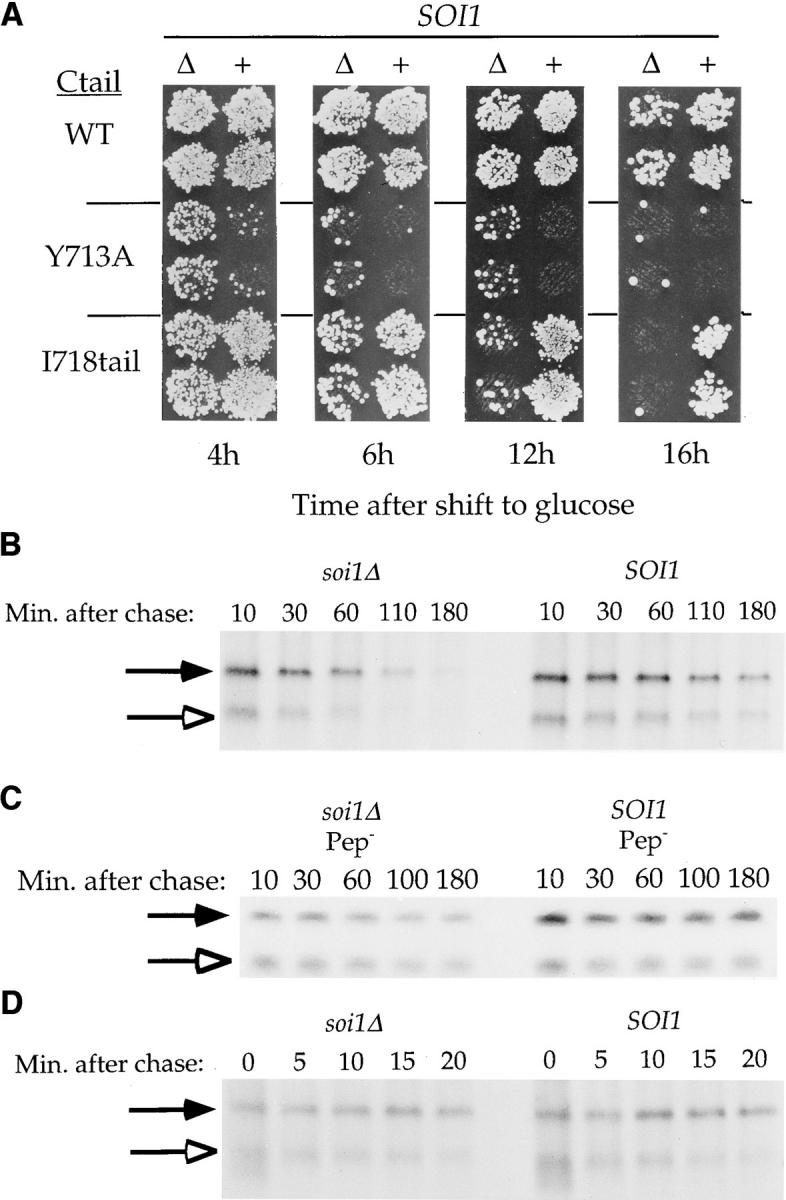
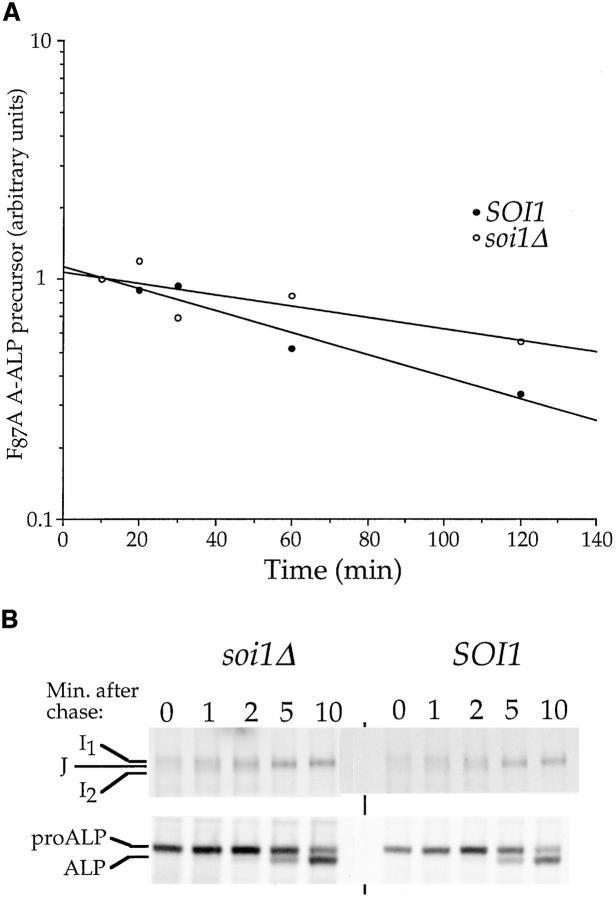
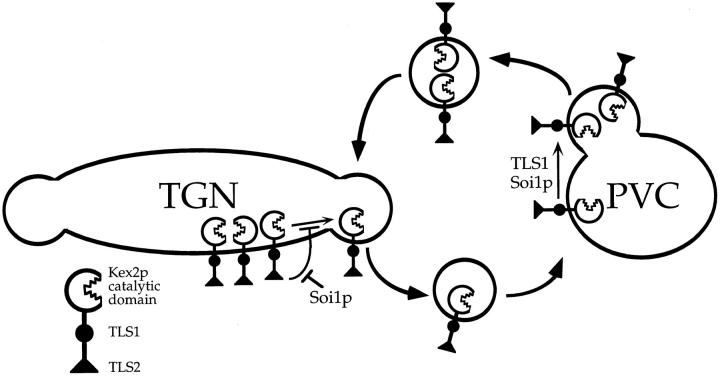
Localization of yeast Kex2 protease to the TGN requires a signal (TLS1) in its cytosolic tail (C-tail). Mutation of TLS1 results in rapid transit of Kex2p to the vacuole. Isolation of suppressors of the Tyr713Ala mutation in TLS1 previously identified three SOI genes. SOI1, cloned by complementation of a sporulation defect, encodes a novel, hydrophilic 3,144-residue protein with homologues in Caenorhabditis elegans, Drosophila melanogaster, and humans. Epitope-tagged Soi1p existed in a detergent-insensitive, sedimentable form. Deletion of SOI1 impaired TGN localization of wild-type Kex2p and a fusion protein containing the C-tail of Ste13p, and also caused missorting of carboxypeptidase Y and accelerated vacuolar degradation of the Vps10p sorting receptor. Deletion of SOI1 improved retention of Tyr713Ala Kex2p in the pro-alpha-factor processing compartment but, unlike the original soi1 alleles, did not increase the half-life of Tyr713Ala Kex2p. These results suggested that Soi1p functions at two steps in the cycling of Kex2p and other proteins between the TGN and prevacuolar compartment (PVC). This hypothesis was confirmed in several ways. Soi1p was shown to be required for optimal function of TLS1. Suppression of the Tyr713Ala mutation by mutation of SOI1 was shown to be caused by activation of a second signal (TLS2) in the Kex2p C-tail. TLS2 delayed exit of Kex2p from the TGN, whereas TLS1 did not affect this step. We propose that Soi1p promotes cycling of TGN membrane proteins between the TGN and PVC by antagonizing a TGN retention signal (TLS2) and facilitating the function of a retrieval signal (TLS1) that acts at the PVC.
Figures





References
-
- Ausubel, F.M., R. Brent, R.E. Kingston, D.D. Moore, J.G. Seidman, J.A. Smith, and K. Struhl. 1989. Mutagenesis of cloned DNA. In Current Protocols in Molecular Biology. Vol. 1. John Wiley and Sons, New York. 8.0.1–8.5.9.
-
- Bordier C. Phase separation of integral membrane proteins in Triton X-114 solution. J Biol Chem. 1981;256:1604–1607. - PubMed
-
- Brenner C, Bevan A, Fuller RS. One-step site-directed mutagenesis of the Kex2 protease oxyanion hole. Curr Biol. 1993;3:498–506. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
