

Desmin is essential for the tensile strength and integrity of myofibrils but not for myogenic commitment, differentiation, and fusion of skeletal muscle
- PMID: 9314534
- PMCID: PMC2139820
- DOI: 10.1083/jcb.139.1.129
Desmin is essential for the tensile strength and integrity of myofibrils but not for myogenic commitment, differentiation, and fusion of skeletal muscle
Abstract
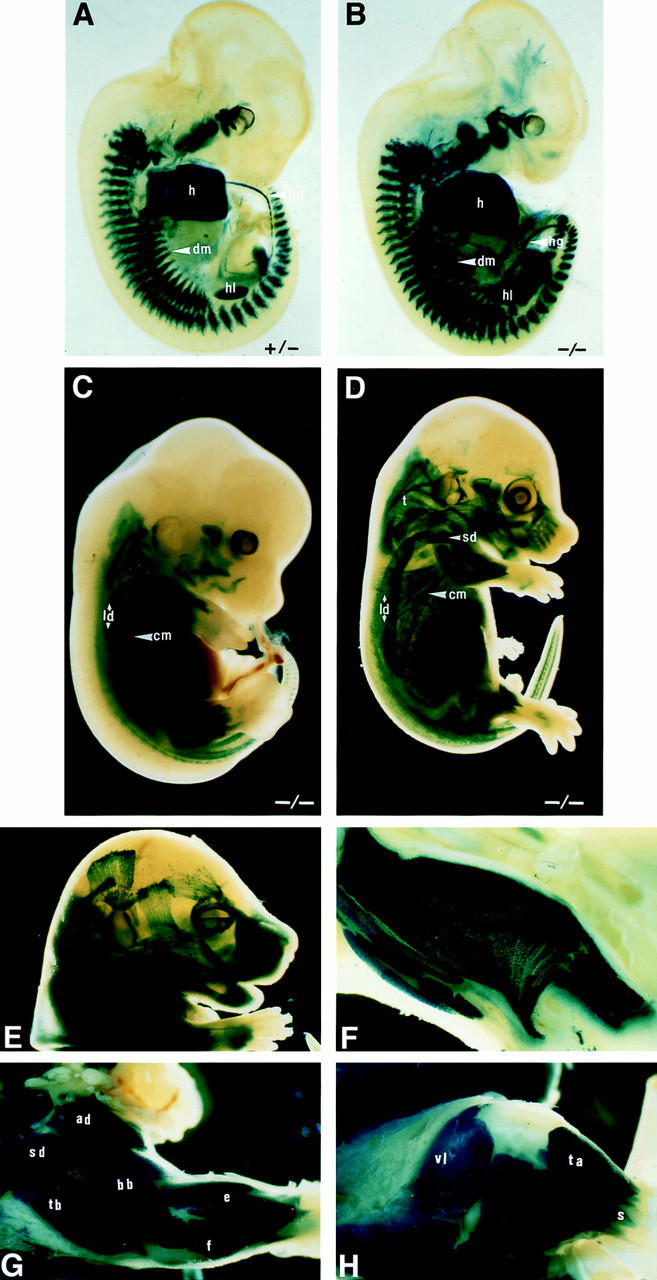
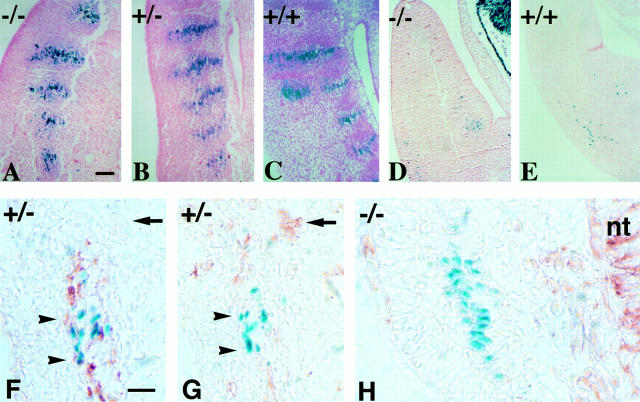
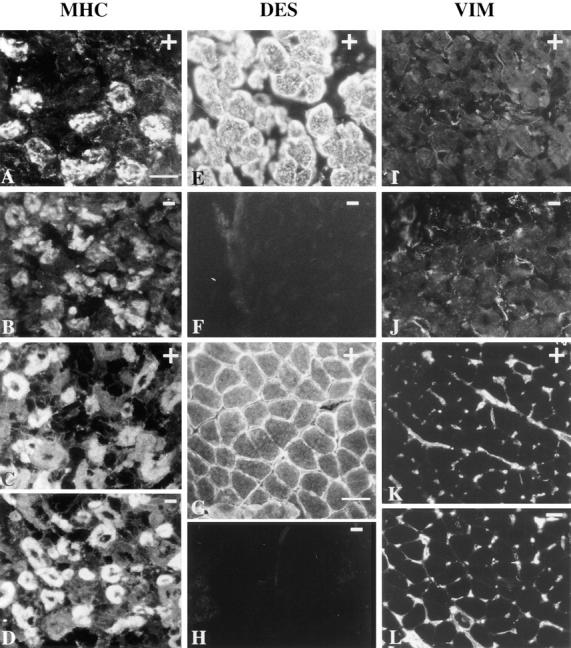
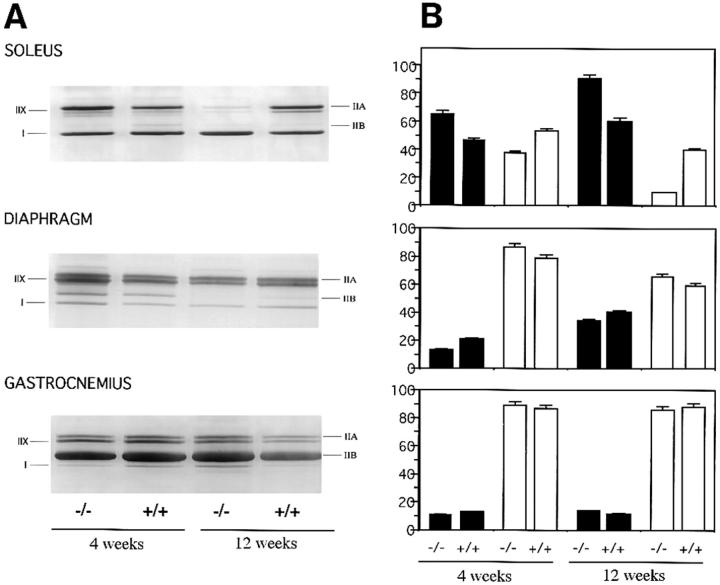
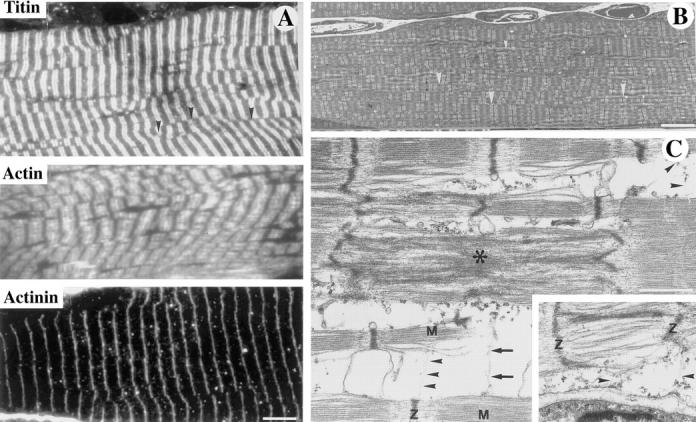
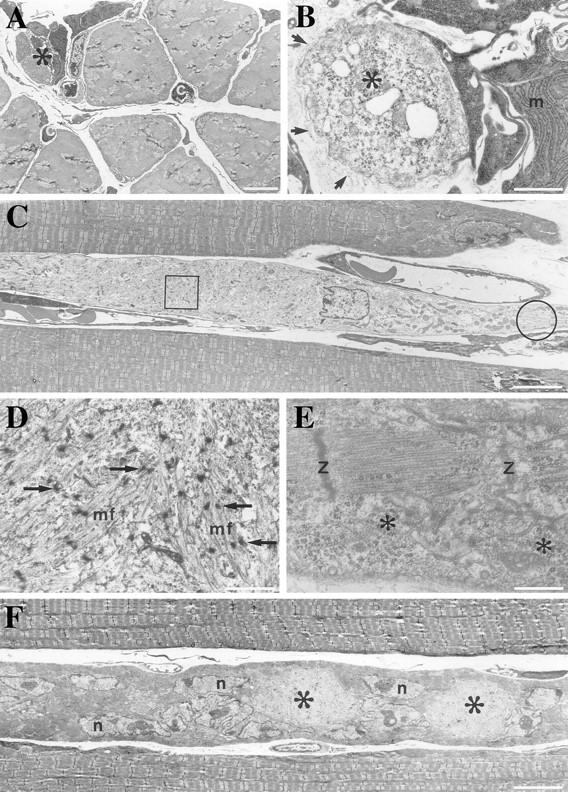
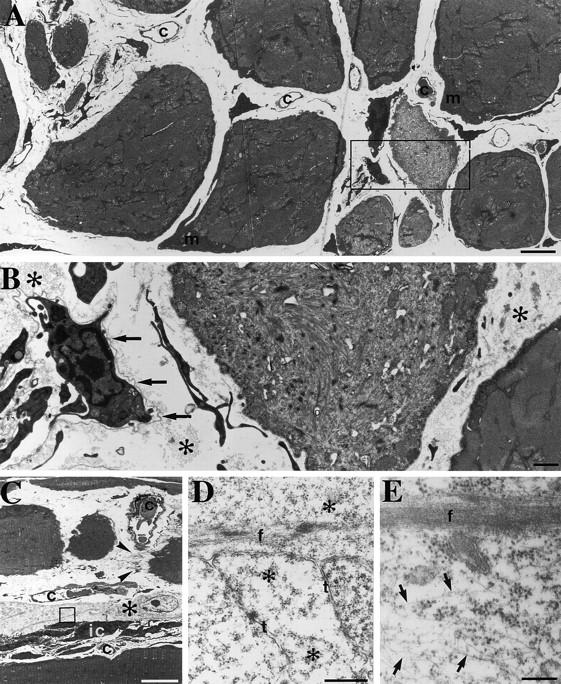
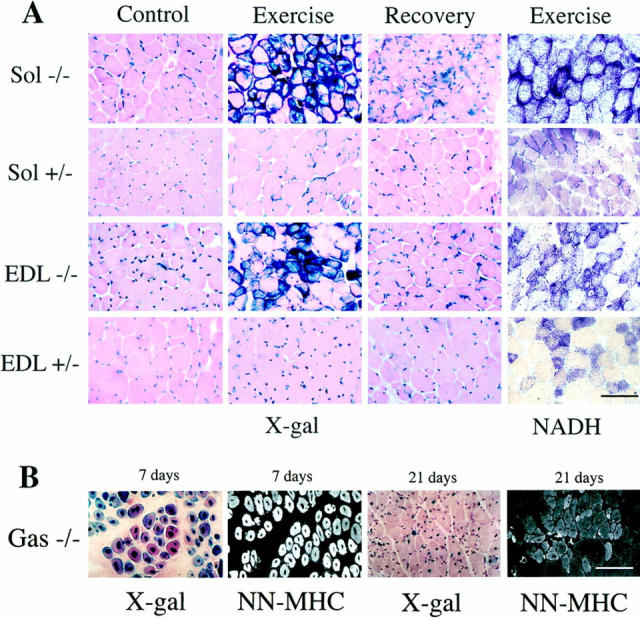
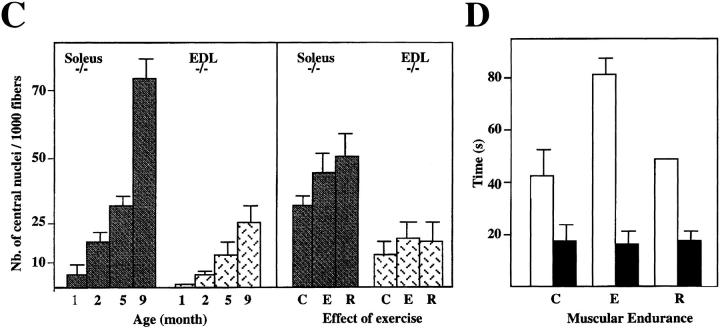
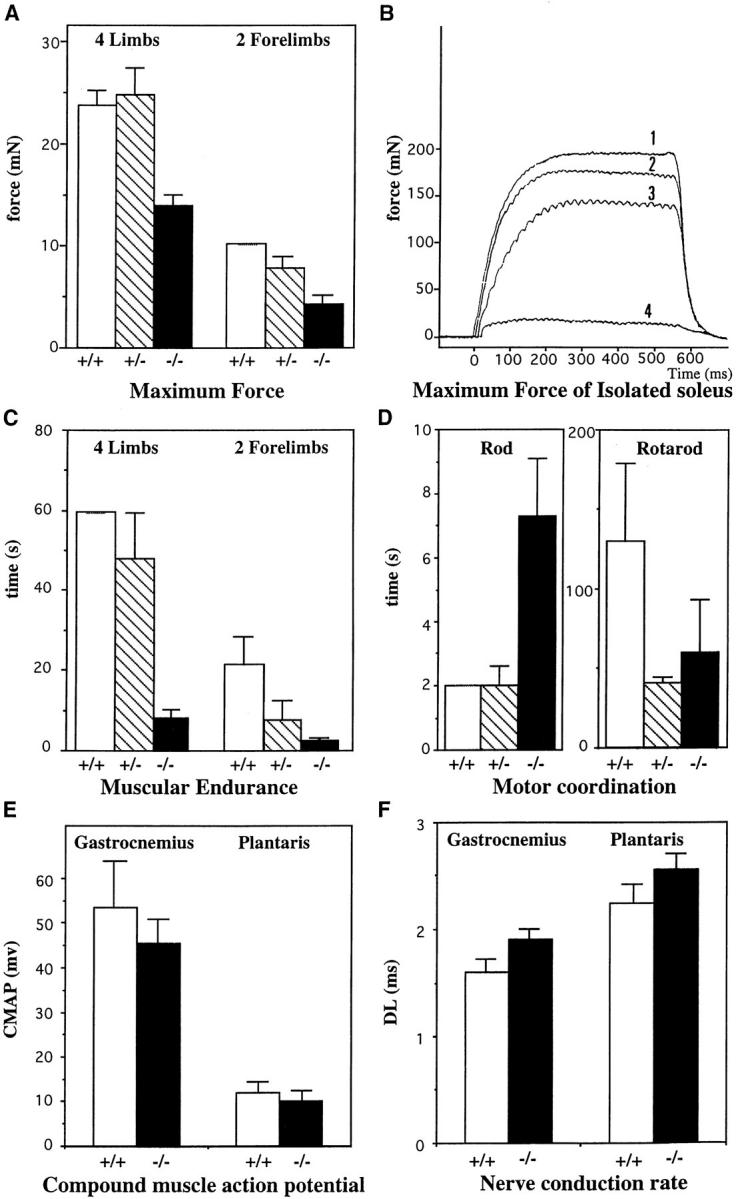
A null mutation was introduced into the mouse desmin gene by homologous recombination. The desmin knockout mice (Des -/-) develop normally and are fertile. However, defects were observed after birth in skeletal, smooth, and cardiac muscles (Li, Z., E. Colucci-Guyon, M. Pincon-Raymond, M. Mericskay, S. Pournin, D. Paulin, and C. Babinet. 1996. Dev. Biol. 175:362-366; Milner, D.J., G. Weitzer, D. Tran, A. Bradley, and Y. Capetanaki. 1996. J. Cell Biol. 134:1255- 1270). In the present study we have carried out a detailed analysis of somitogenesis, muscle formation, maturation, degeneration, and regeneration in Des -/- mice. Our results demonstrate that all early stages of muscle differentiation and cell fusion occur normally. However, after birth, modifications were observed essentially in weight-bearing muscles such as the soleus or continually used muscles such as the diaphragm and the heart. In the absence of desmin, mice were weaker and fatigued more easily. The lack of desmin renders these fibers more susceptible to damage during contraction. We observed a process of degeneration of myofibers, accompanied by macrophage infiltration, and followed by a process of regeneration. These cycles of degeneration and regeneration resulted in a relative increase in slow myosin heavy chain (MHC) and decrease in fast MHC. Interestingly, this second wave of myofibrillogenesis during regeneration was often aberrant and showed signs of disorganization. Subsarcolemmal accumulation of mitochondria were also observed in these muscles. The lack of desmin was not compensated by an upregulation of vimentin in these mice either during development or regeneration. Absence of desmin filaments within the sarcomere does not interfere with primary muscle formation or regeneration. However, myofibrillogenesis in regenerating fibers is often abortive, indicating that desmin may be implicated in this repair process. The results presented here show that desmin is essential to maintain the structural integrity of highly solicited skeletal muscle.
Figures
References
-
- Agbulut O, Li Z, Mouly V, Butler-Browne GS. Analysis of skeletal and cardiac muscle from desmin knock-out and normal mice by high resolution separation of MHC isoforms. Biol Cell. 1997;88:131–135. - PubMed
-
- Allen RE, Rankin LL, Greene EA, Boxhorn LK, Johnson SE, Taylor RG, Pierce PA. Desmin is present in proliferating rat muscle satellite cells but not in bovine muscle satellite cells. J Cell Physiol. 1991;149:525–535. - PubMed
-
- Ariza A, Coll J, Fernandez-Figueras MT, Lopez MD, Mate JL, Garcia O, Fernandez-Vasalo A, Navas-Palacios JJ. Desmin myopathy: a multisystem disorder involving skeletal, cardiac, and smooth muscle. J Hum Pathol. 1995;26:1032–1037. - PubMed
-
- Babai F, Musevi-Aghdam J, Schurch W, Royal A, Gabbiani G. Coexpression of α-sarcomeric actin, α-smooth muscle actin and desmin during myogenesis in rat and mouse embryos. I. Skeletal muscle. Differentiation. 1990;44:132–142. - PubMed
-
- Baribault H, Price J, Miyai K, Oshima RG. Mid-gestational lethality in mice lacking keratin 8. Genes Dev. 1993;7:1191–1202. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
