

Bax deletion further orders the cell death pathway in cerebellar granule cells and suggests a caspase-independent pathway to cell death
- PMID: 9314540
- PMCID: PMC2139809
- DOI: 10.1083/jcb.139.1.205
Bax deletion further orders the cell death pathway in cerebellar granule cells and suggests a caspase-independent pathway to cell death
Abstract
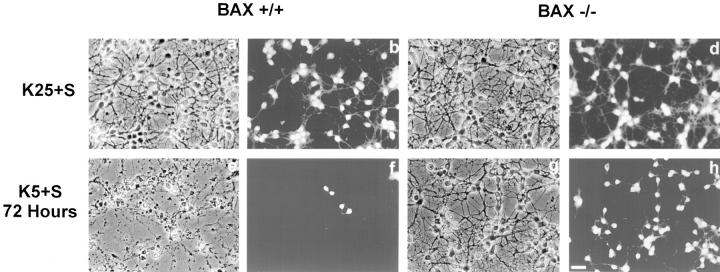
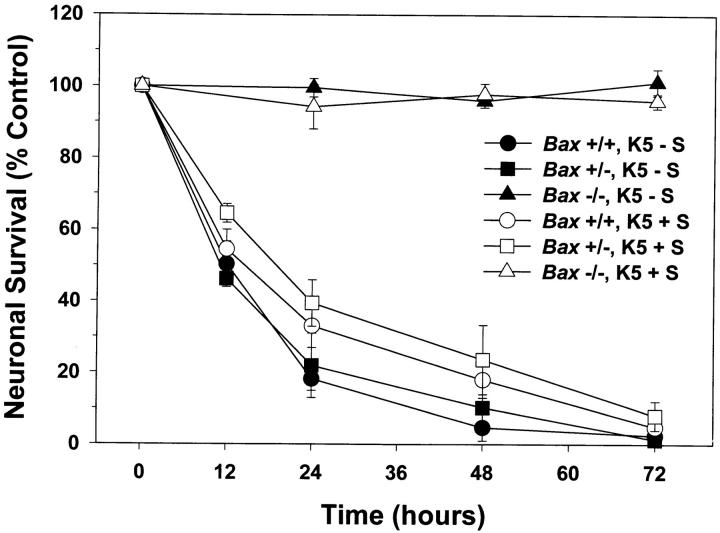
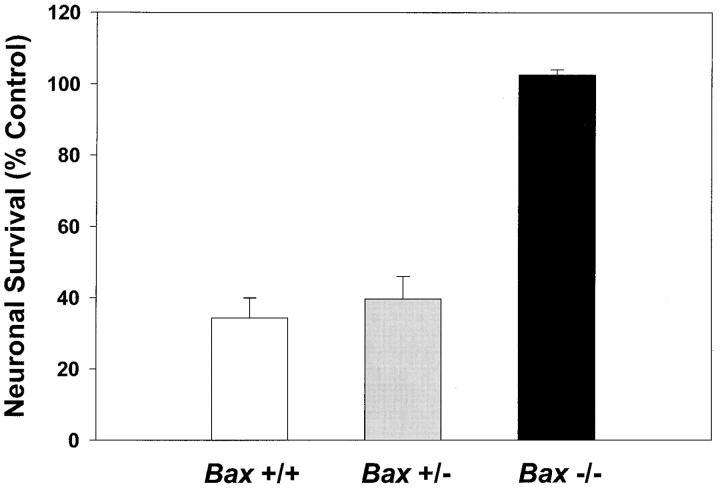
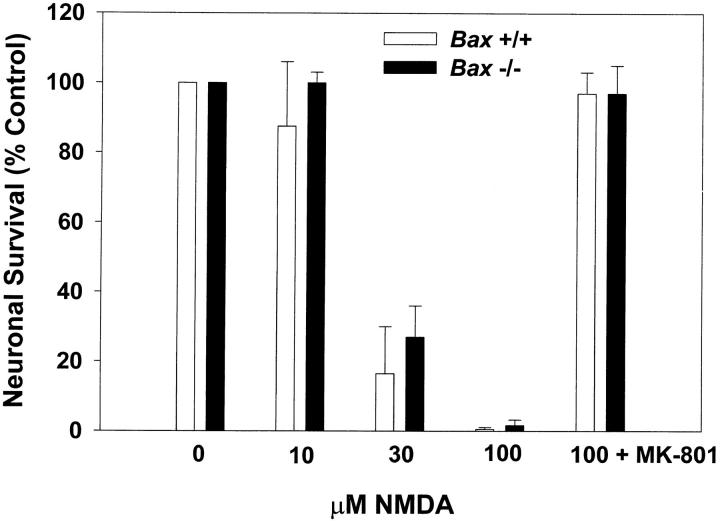
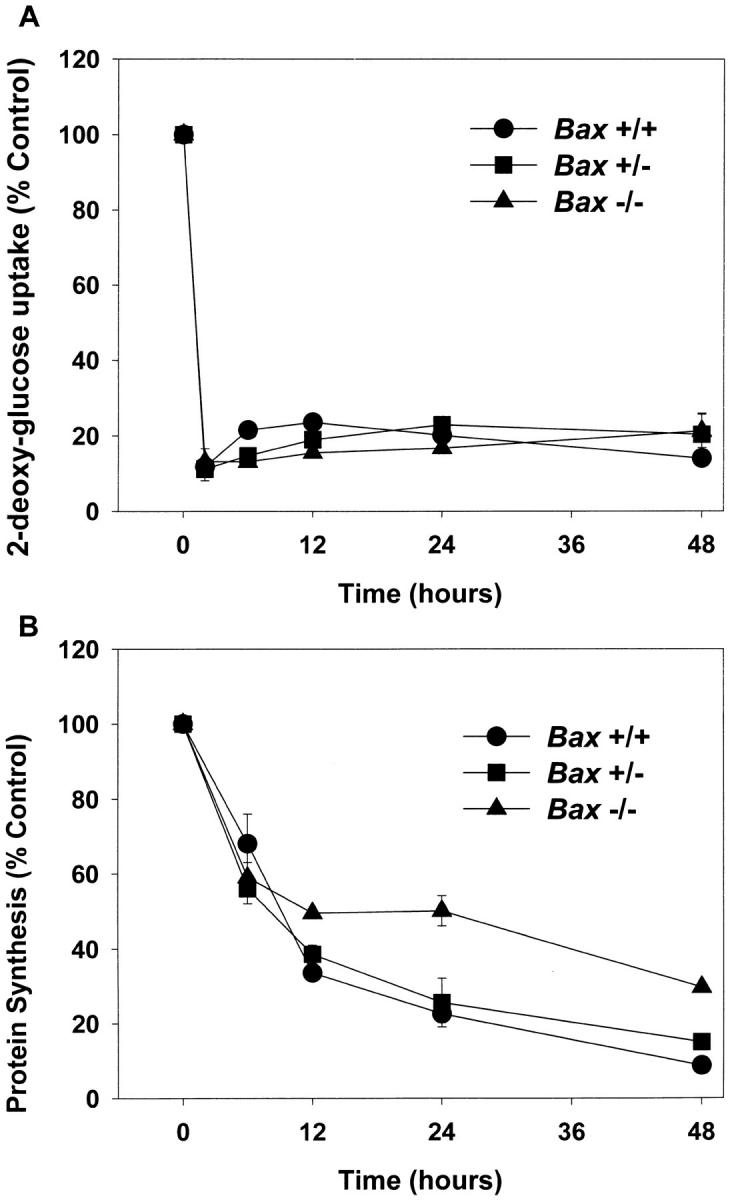
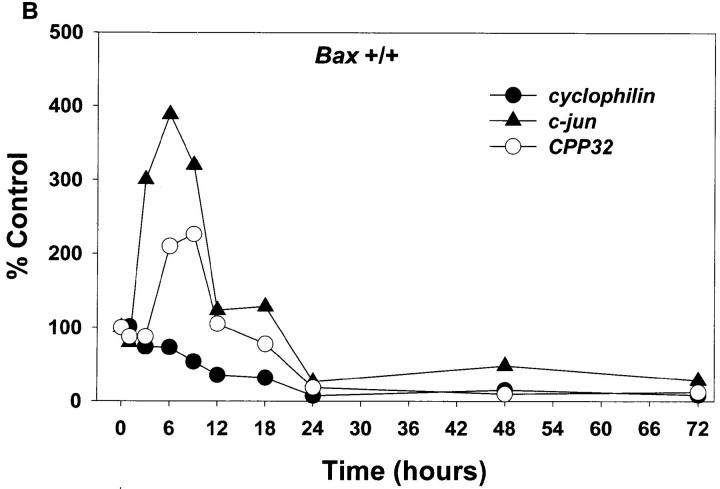
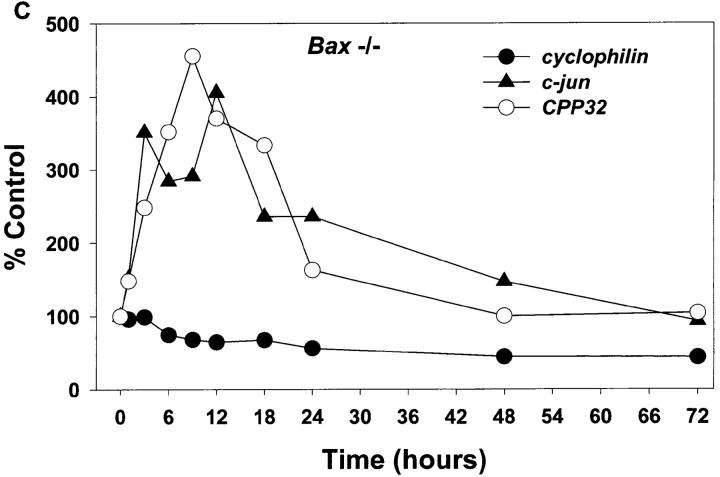
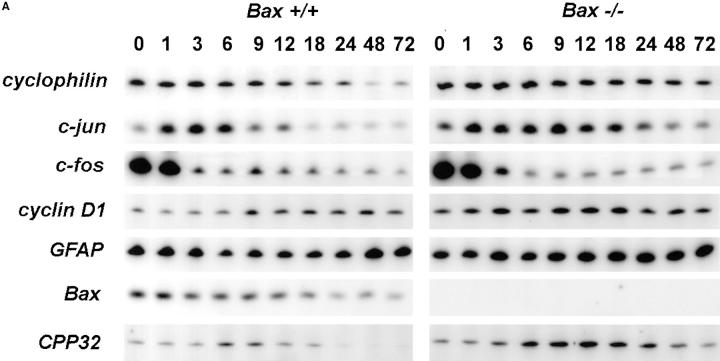
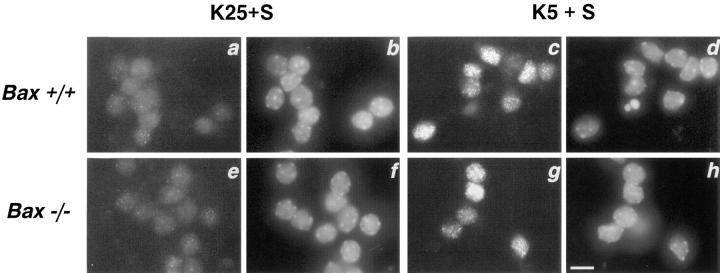
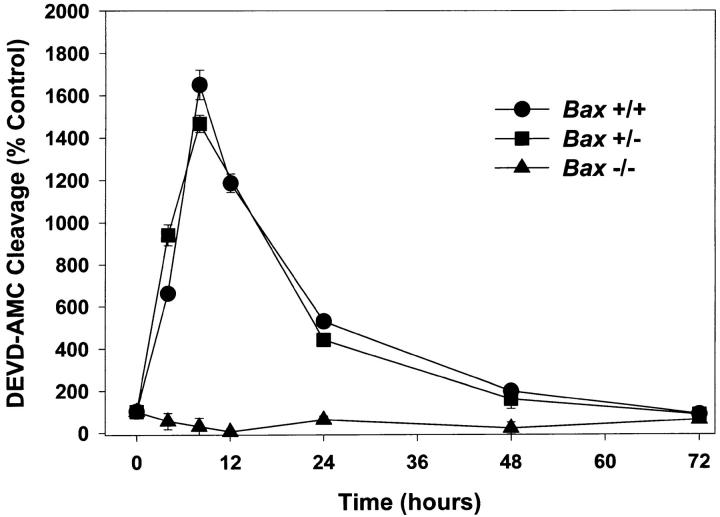
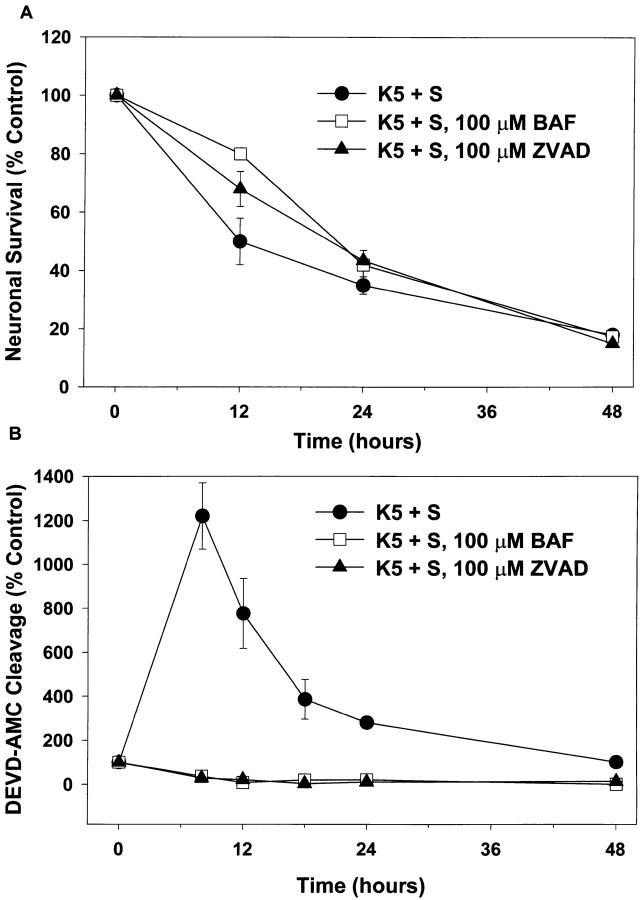
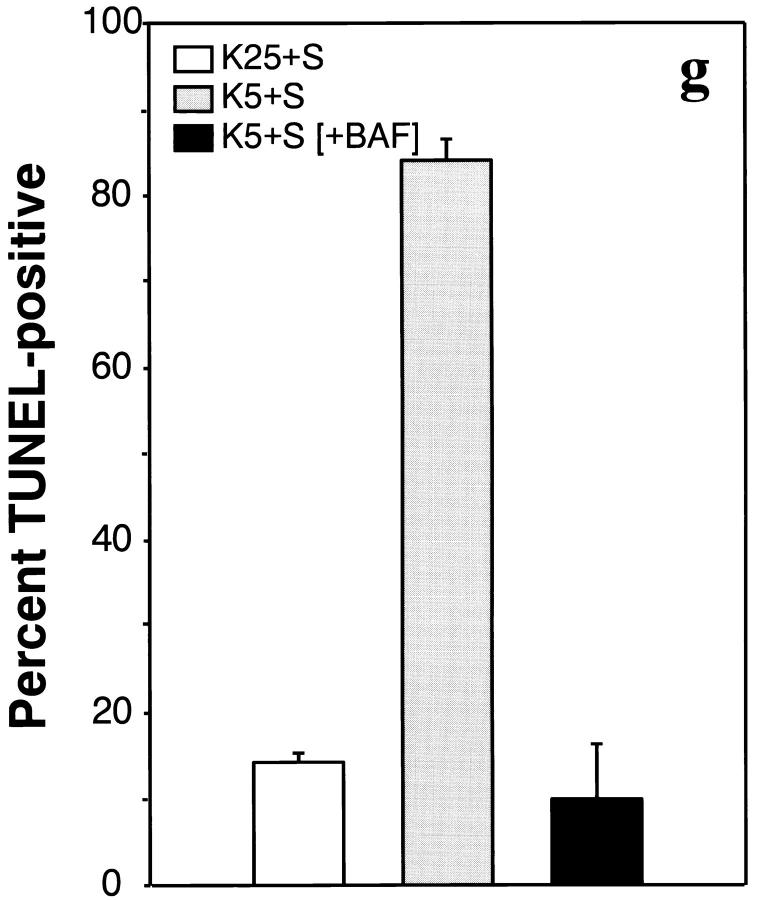
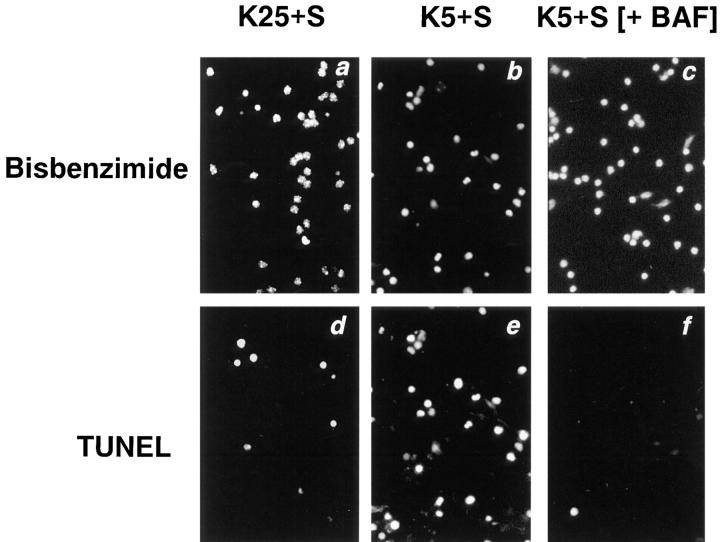
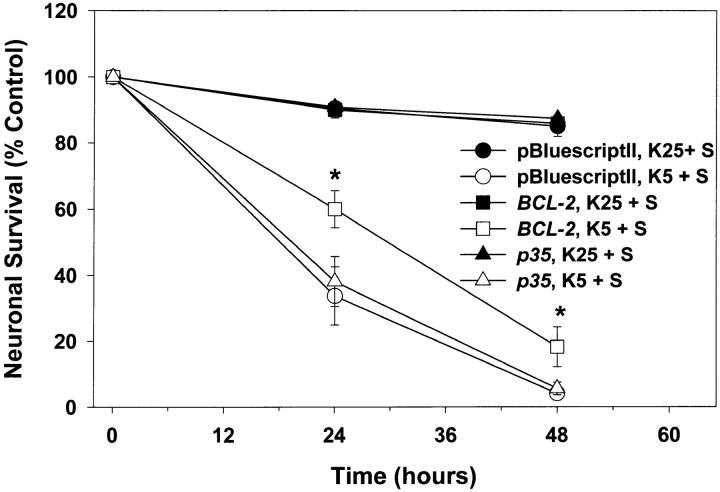
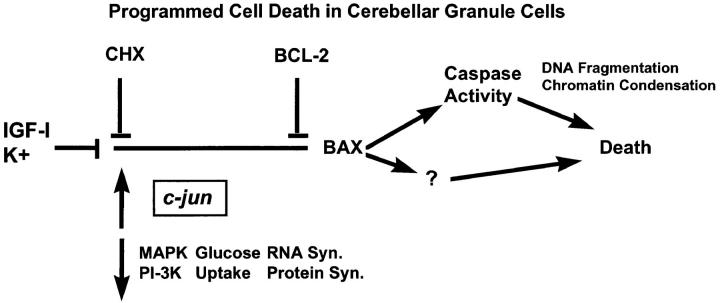
Dissociated cerebellar granule cells maintained in medium containing 25 mM potassium undergo an apoptotic death when switched to medium with 5 mM potassium. Granule cells from mice in which Bax, a proapoptotic Bcl-2 family member, had been deleted, did not undergo apoptosis in 5 mM potassium, yet did undergo an excitotoxic cell death in response to stimulation with 30 or 100 microM NMDA. Within 2 h after switching to 5 mM K+, both wild-type and Bax-deficient granule cells decreased glucose uptake to <20% of control. Protein synthesis also decreased rapidly in both wild-type and Bax-deficient granule cells to 50% of control within 12 h after switching to 5 mM potassium. Both wild-type and Bax -/- neurons increased mRNA levels of c-jun, and caspase 3 (CPP32) and increased phosphorylation of the transactivation domain of c-Jun after K+ deprivation. Wild-type granule cells in 5 mM K+ increased cleavage of DEVD-aminomethylcoumarin (DEVD-AMC), a fluorogenic substrate for caspases 2, 3, and 7; in contrast, Bax-deficient granule cells did not cleave DEVD-AMC. These results place BAX downstream of metabolic changes, changes in mRNA levels, and increased phosphorylation of c-Jun, yet upstream of the activation of caspases and indicate that BAX is required for apoptotic, but not excitotoxic, cell death. In wild-type cells, Boc-Asp-FMK and ZVAD-FMK, general inhibitors of caspases, blocked cleavage of DEVD-AMC and blocked the increase in TdT-mediated dUTP nick end labeling (TUNEL) positivity. However, these inhibitors had only a marginal effect on preventing cell death, suggesting a caspase-independent death pathway downstream of BAX in cerebellar granule cells.
Figures
References
-
- Armstrong RC, Aja T, Xiang J, Gaur S, Krebs JF, Hoang K, Bai X, Korsmeyer SJ, Karanewsky DS, Fritz LC, Tomaselli KJ. Fas-induced activation of the cell death-related protease CPP32 is inhibited by Bcl-2 and by ICE family protease inhibitors. J Biol Chem. 1996;271:16850–16855. - PubMed
-
- Asahi M, Hoshimaru M, Uemura Y, Tokime T, Kojima M, Ohtsuka T, Matsuura N, Aoki T, Shibahara K, Kikuchi H. Expression of interleukin-1β converting enzyme gene family and bcl-2 gene family in the rat brain following permanent occlusion of the middle cerebral artery. J Cerebr Blood Flow Metab. 1997;17:11–18. - PubMed
-
- Balazs R, Jorgensen OS, Hack N. N-Methyl-D-Aspartate promotes the survival of cerebellar granule cells in culture. Neuroscience. 1988;27:437–451. - PubMed
-
- Binetruy B, Smeal T, Karin M. HaRas augments c-Jun activity and stimulates phosphorylation of its activation domain. Nature (Lond) 1991;351:122–127. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
