

How many thymocytes audition for selection?
- PMID: 9314563
- PMCID: PMC2199071
- DOI: 10.1084/jem.186.7.1149
How many thymocytes audition for selection?
Abstract
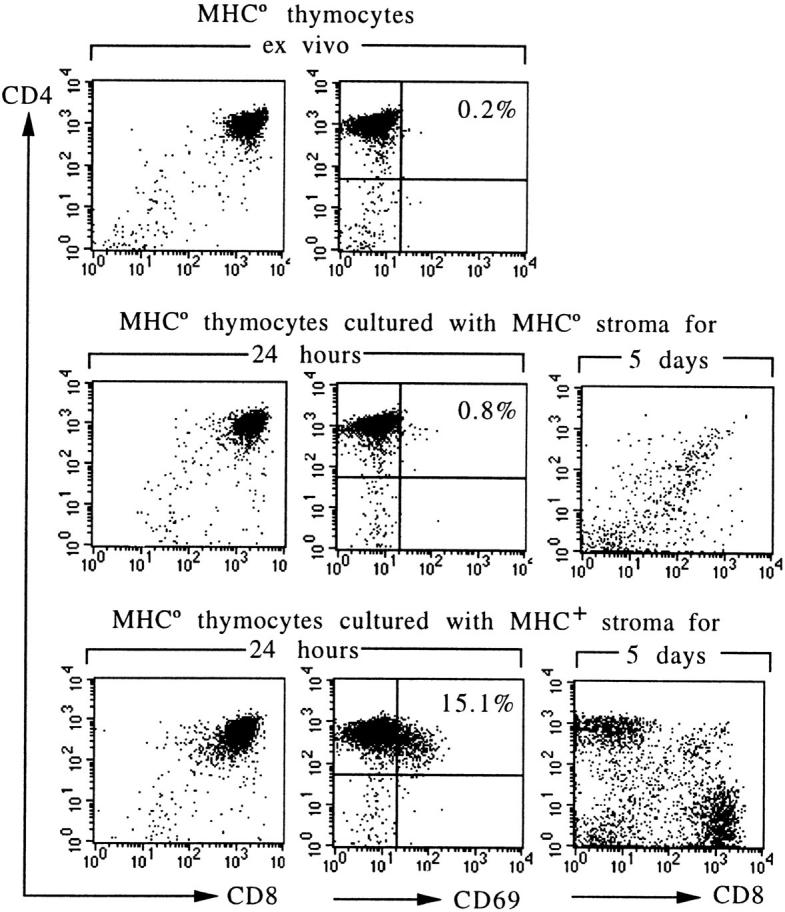
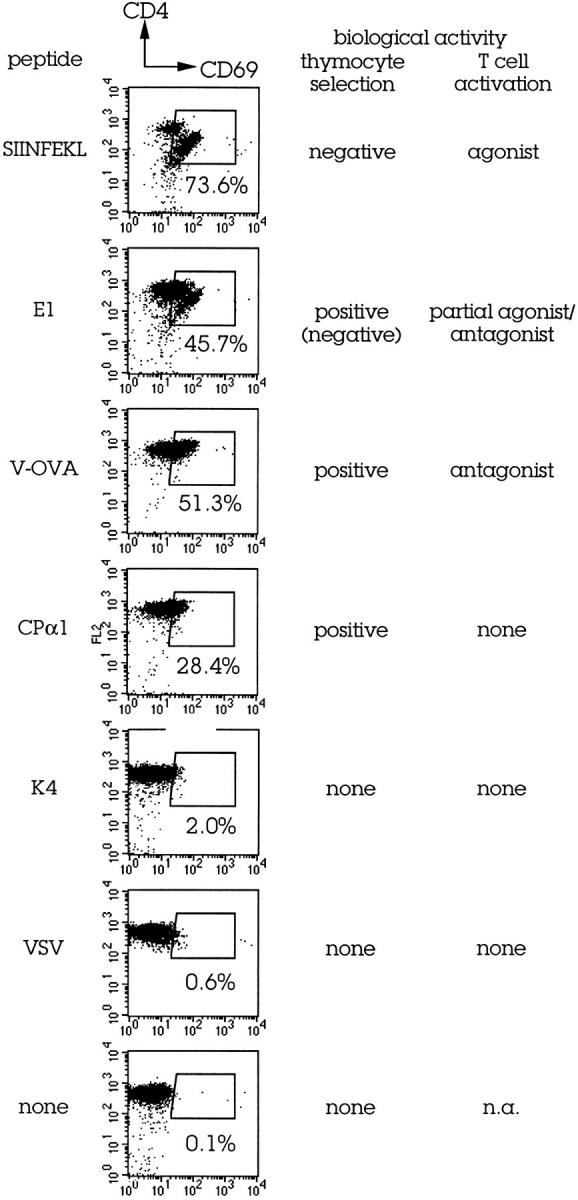
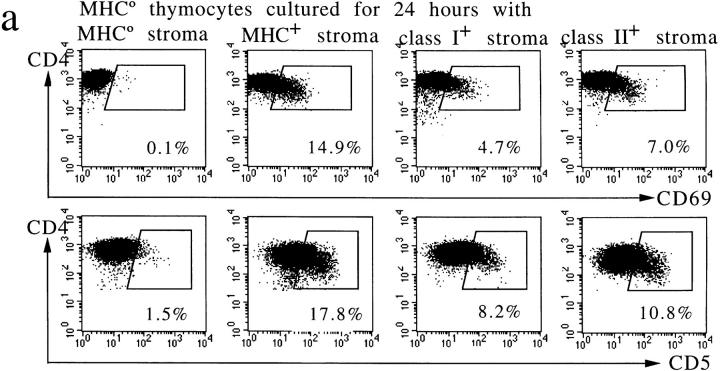
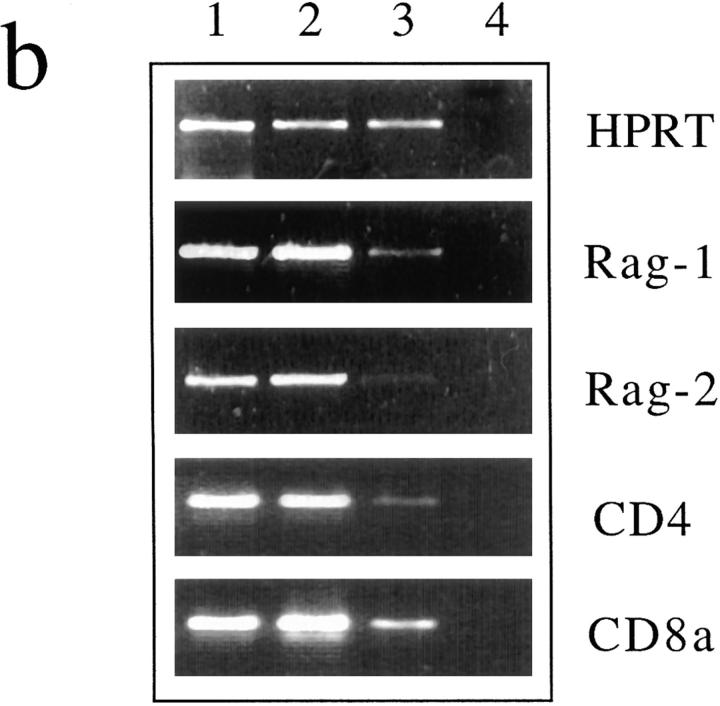
T cell maturation requires the rearrangement of clonotypic T cell receptors (TCR) capable of interacting with major histocompatibility complex (MHC) ligands to initiate positive and negative selection. Only 3-5% of thymocytes mature to join the peripheral T cell pool. To investigate the basis for this low success rate, we have measured the frequency of preselection thymocytes capable of responding to MHC. As many as one in five MHC-naive thymocytes show upregulation of activation markers on exposure to MHC-expressing thymic stroma in short-term reaggregate culture. The majority of these cells display physiological changes consistent with entry into the selection process within 24 h. By exposing TCR transgenic thymocytes to a range of MHC-peptide complexes, we show that CD69 induction is indicative of thymocyte selection, positive or negative. Our data provide evidence that the fraction of thymocytes that qualify to enter the thymic selection process far exceeds the fraction that successfully complete it, and suggest that most MHC-reactive thymocytes are actively eliminated in the course of selection.
Figures
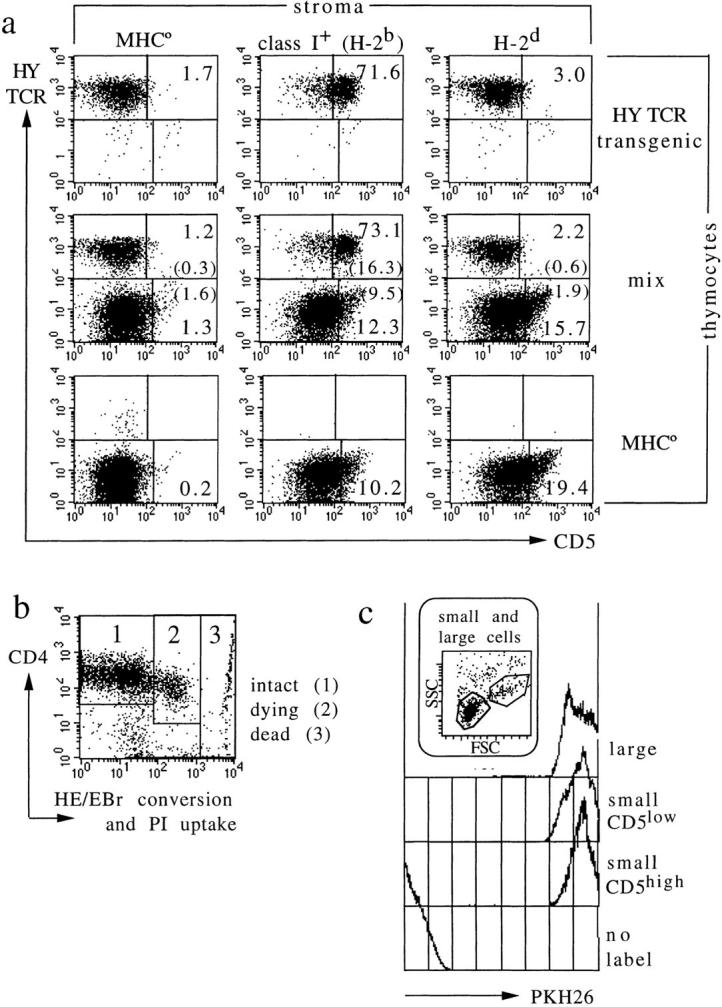
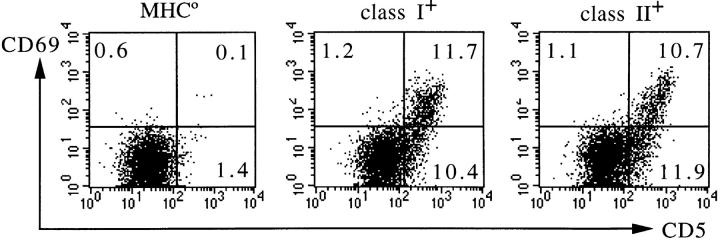
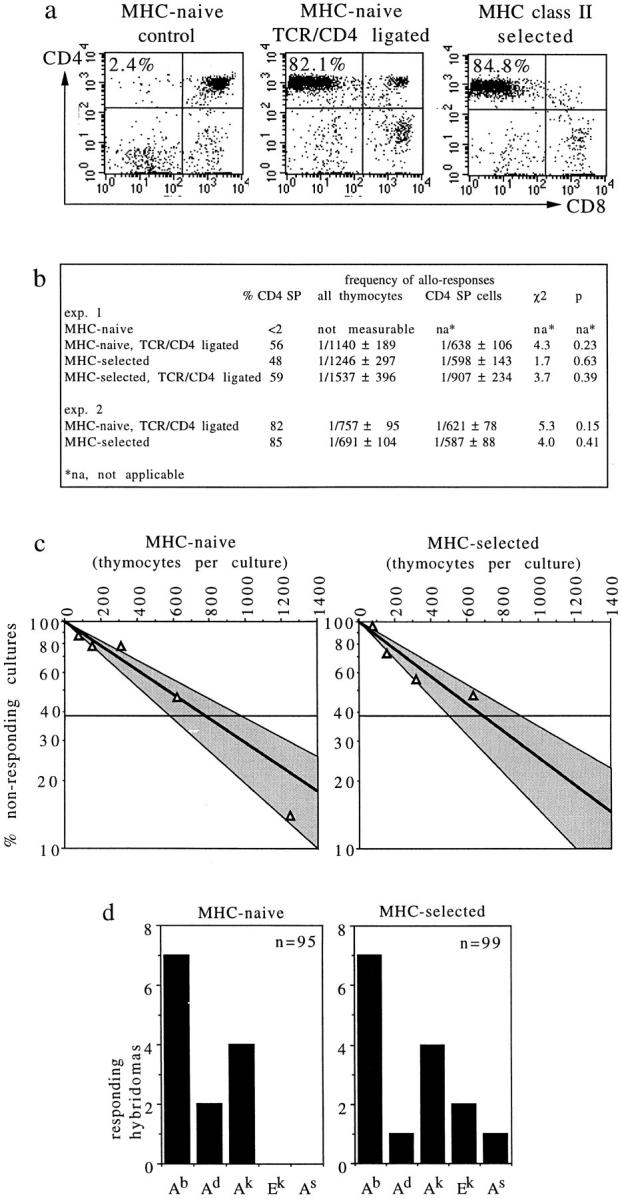
References
-
- Zinkernagel RM, Doherty PC. MHC-restricted cytotoxic T cells: studies on the biological role of polymorphic major transplantation antigens determining T-cell restriction-specificity, function, and responsiveness. Adv Immunol. 1979;27:51–177. - PubMed
-
- Bevan MJ. In a radiation chimaera, host H-2 antigens determine immune responsiveness of donor cytotoxic cells. Nature (Lond) 1977;269:417–418. - PubMed
-
- Jorgensen JL, Reay PA, Ehrich EW, Davis MM. Molecular components of T-cell recognition. Annu Rev Immunol. 1992;10:835–873. - PubMed
-
- Kisielow P, von Boehmer H. Development and selection of T cells: facts and puzzles. Adv Immunol. 1995;58:87–209. - PubMed
-
- Gilfillan S, Benoist C, Mathis D. Mice lacking terminal deoxynucleotidyl transferase: adult mice with a fetal antigen receptor repertoire. Immunol Rev. 1995;148:201–219. - PubMed
