

Late compartments of amyloid precursor protein transport in SY5Y cells are involved in beta-amyloid secretion
- PMID: 9315893
- PMCID: PMC6793894
- DOI: 10.1523/JNEUROSCI.17-20-07714.1997
Late compartments of amyloid precursor protein transport in SY5Y cells are involved in beta-amyloid secretion
Abstract
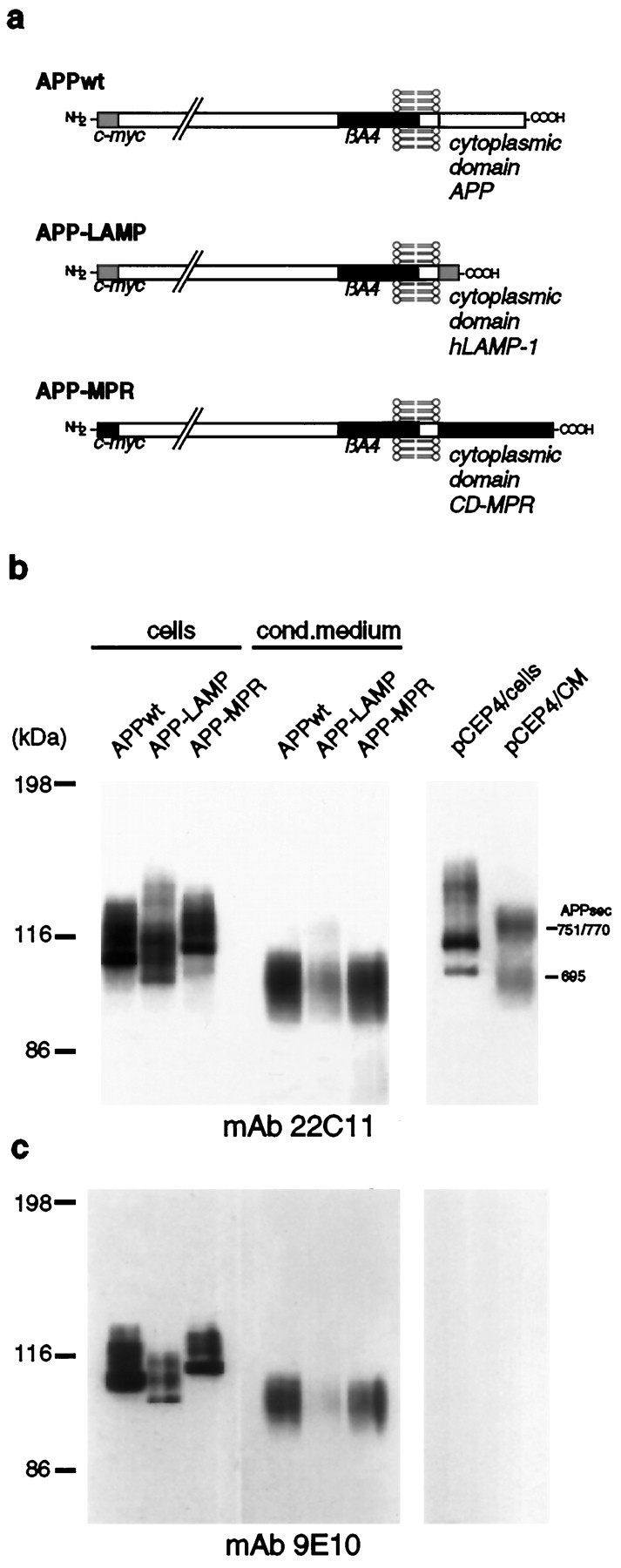
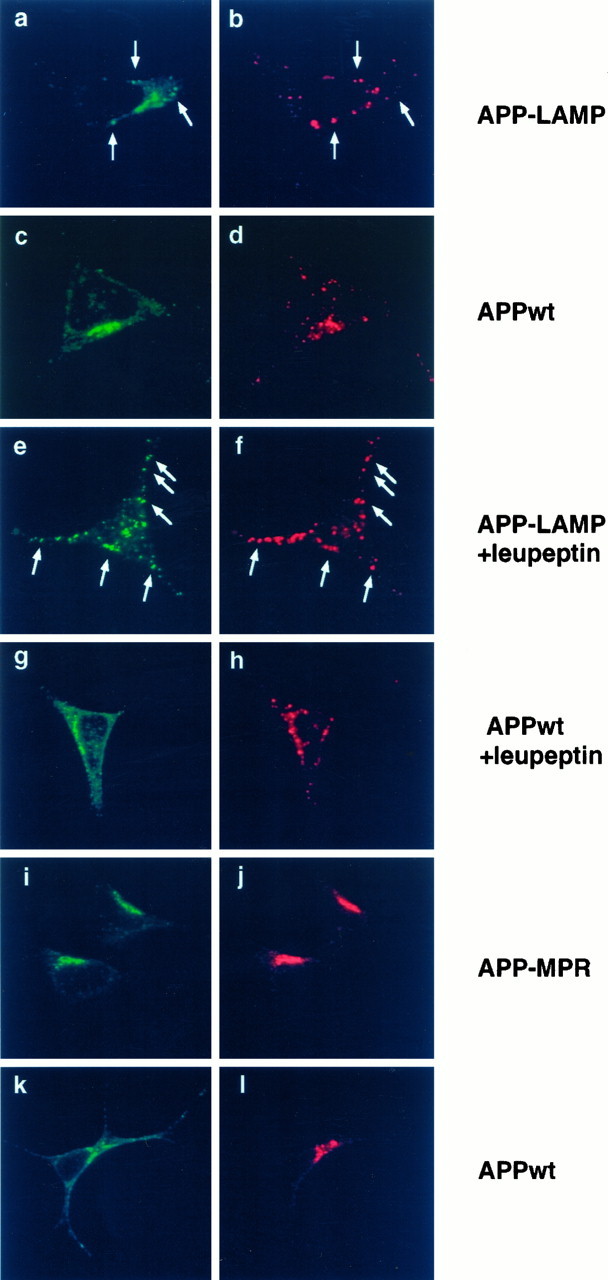
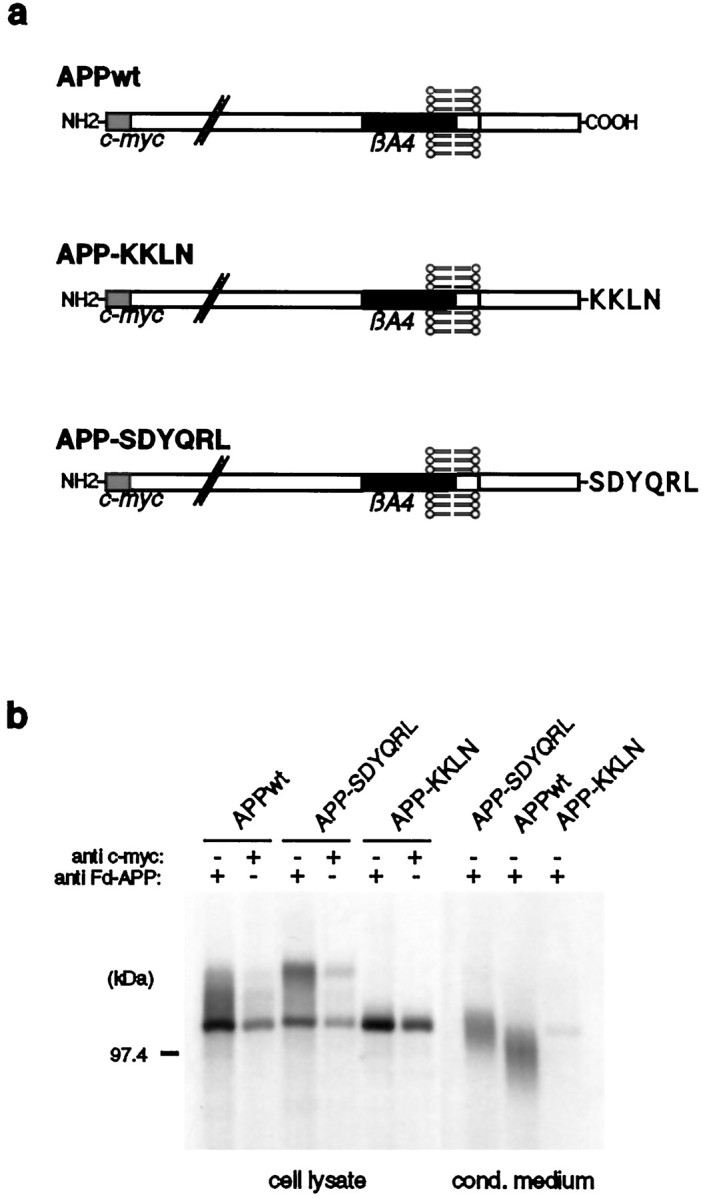
Amyloid plaques, composed mainly of the 39-43 amino acid betaA4 peptide, are a characteristic feature of Alzheimer's disease. Generation of betaA4 by proteolytic processing of the amyloid precursor protein (APP) is thought to occur in a pathway that includes the activity of two as yet unknown proteases, with beta-secretase cleaving at the N terminus and gamma-secretase releasing the C terminus of betaA4. Inhibition studies and the finding that cell surface APP can serve as a direct precursor of betaA4 suggest that the endosomal/lysosomal compartment is involved in the proteolysis of APP into betaA4. In this study we targeted APP695 chimeric proteins directly into the endosomal/lysosomal compartment. This decreased the amount of released betaA4, while the generation of the betaA4 N terminus continued. APP695 proteins were constructed also, which carried sorting signals responsible for recycling between the trans-Golgi network (TGN) and the cell surface. These proteins were processed into secreted betaA4 at even higher levels than wild-type APP695. Moreover, retention of APP695 proteins in the endoplasmic reticulum led to neither betaA4 secretion nor to processing by beta-secretase in human SH-SY5Y neuroblastoma cells. These data suggest that a beta-cleavage activity resides in a late endosomal compartment and that a gamma-cleavage occurs in early endosomes, resulting in the generation of betaA4 peptides with the majority ending at residue 40.
Figures





References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous