

Aggregated amyloid-beta protein induces cortical neuronal apoptosis and concomitant "apoptotic" pattern of gene induction
- PMID: 9315895
- PMCID: PMC6793913
- DOI: 10.1523/JNEUROSCI.17-20-07736.1997
Aggregated amyloid-beta protein induces cortical neuronal apoptosis and concomitant "apoptotic" pattern of gene induction
Abstract
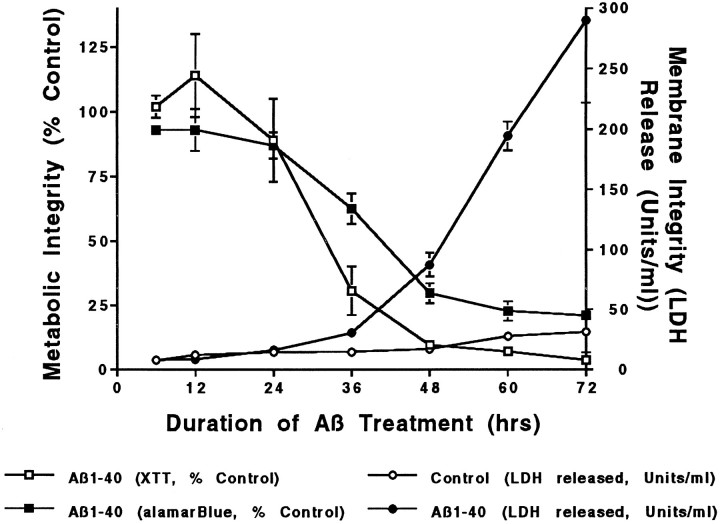
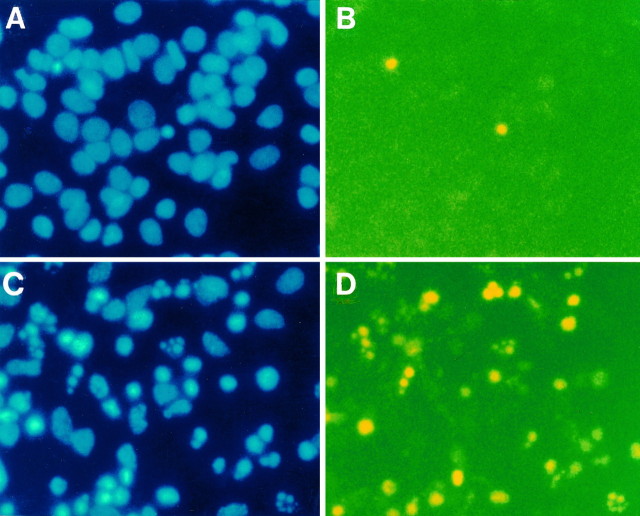
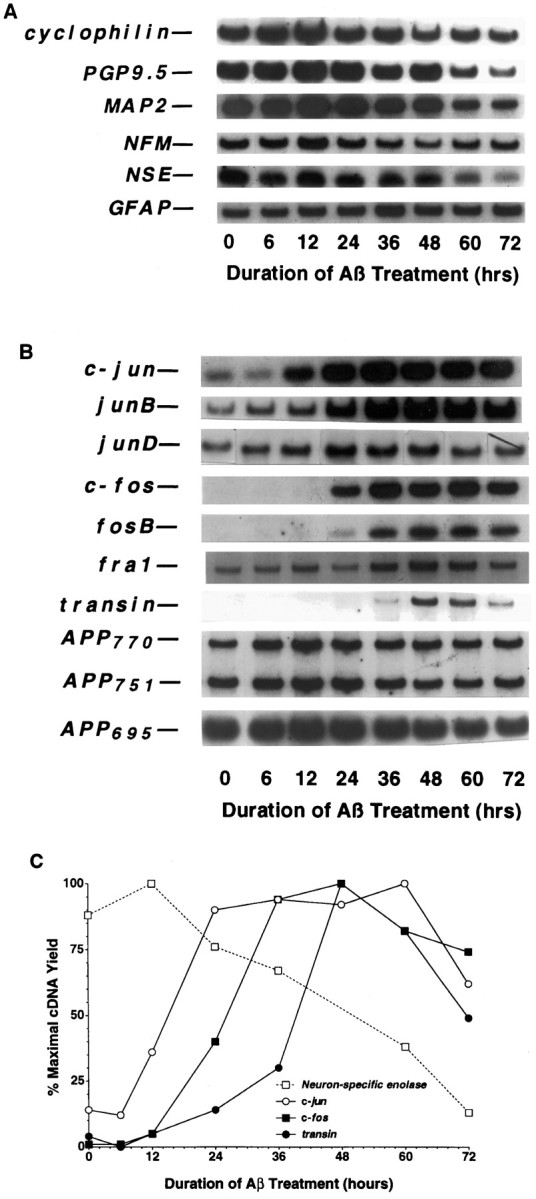
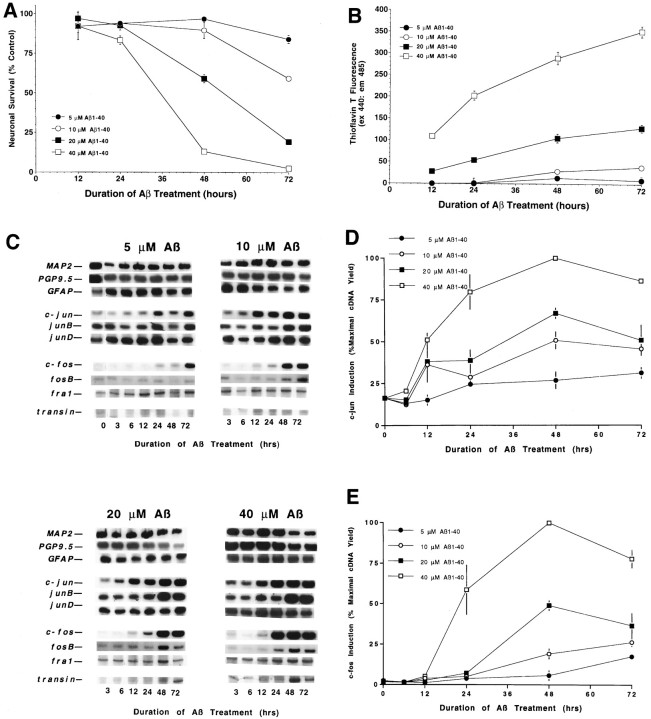
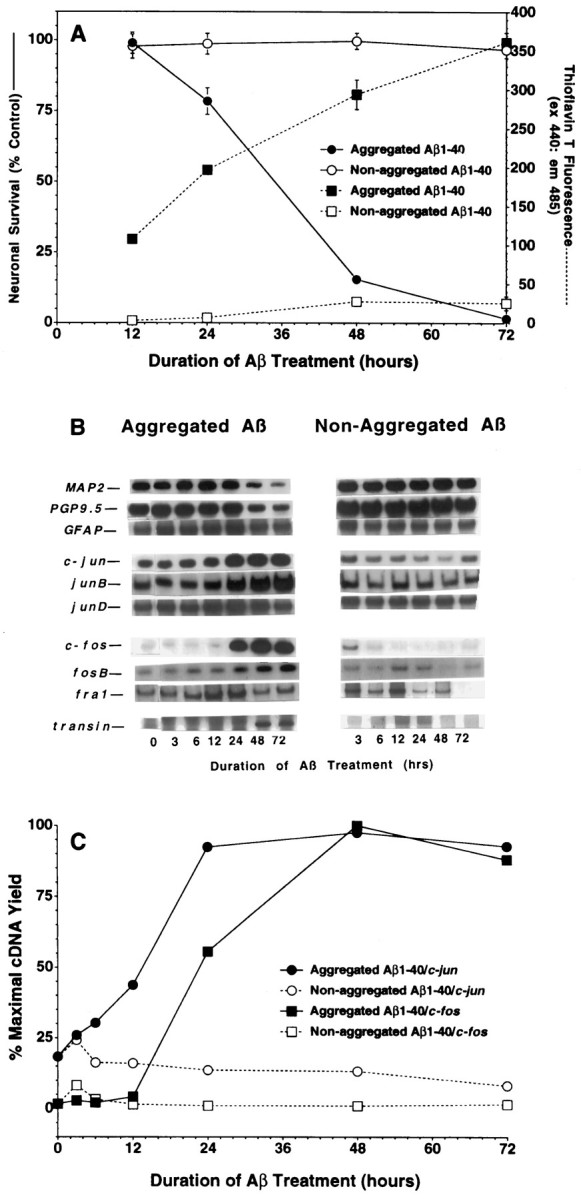
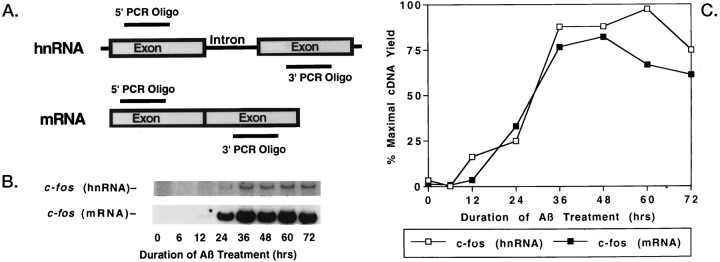
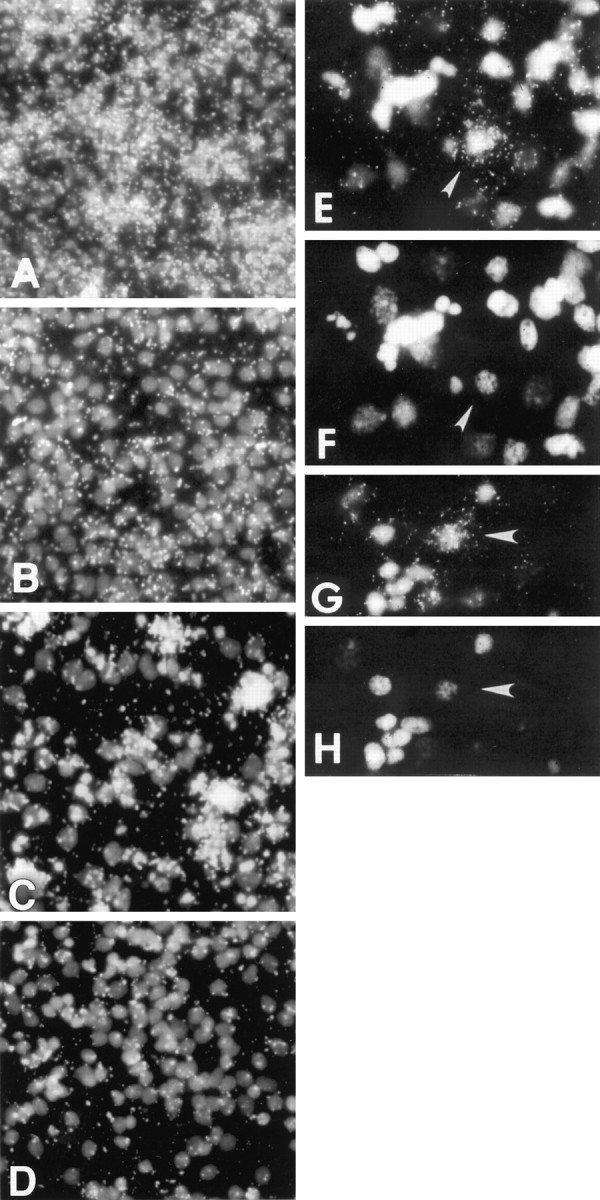
To gain a molecular understanding of neuronal responses to amyloid-beta peptide (Abeta), we have analyzed the effects of Abeta treatment on neuronal gene expression in vitro by quantitative reverse transcription-PCR and in situ hybridization. Treatment of cultured rat cortical neurons with Abeta1-40 results in a widespread apoptotic neuronal death. Associated with death is an induction of several members of the immediate early gene family. Specifically, we (1) report the time-dependent and robust induction of c-jun, junB, c-fos, and fosB, as well as transin, which is induced by c-Jun/c-Fos heterodimers and encodes an extracellular matrix protease; these gene inductions appear to be selective because other Jun and Fos family members, i.e., junD and fra-1, are induced only marginally; (2) show that the c-jun induction is widespread, whereas c-fos expression is restricted to a subset of neurons, typically those with condensed chromatin, which is a hallmark of apoptosis; (3) correlate gene induction and neuronal death by showing that each has a similar dose-response to Abeta; and (4) demonstrate that both cell death and immediate early gene induction are dependent on Abeta aggregation state. This overall gene expression pattern during this "physiologically inappropriate" apoptotic stimulus is markedly similar to the pattern we previously identified after a "physiologically appropriate" stimulus, i.e., the NGF deprivation-induced death of sympathetic neurons. Hence, the parallels identified here further our understanding of the genetic alterations that may lead neurons to apoptosis in response to markedly different insults.
Figures
References
-
- Anderson AJ, Cummings BJ, Cotman CW. Increased immunoreactivity for Jun- and Fos-related proteins in Alzheimer’s disease: association with pathology. Exp Neurol. 1994;125:286–295. - PubMed
-
- Anderson AJ, Pike CJ, Cotman CW. Differential induction of immediate early gene proteins in cultured neurons by β-amyloid (Aβ): association of c-jun with Aβ-induced apoptosis. J Neurochem. 1995;65:1487–1498. - PubMed
-
- Angel P, Karin M. The role of Jun, Fos, and the Ap-1 complex in cell proliferation and transformation. Biochim Biophys Acta. 1991;1072:129–157. - PubMed
-
- Behl C, Davis JB, Klier FG, Schubert D. Amyloid beta peptide induces necrosis rather than apoptosis. Brain Res. 1994a;645:254–264. - PubMed
-
- Behl C, Davis JB, Lesley R, Schubert D. Hydrogen peroxide mediates amyloid beta protein toxicity. Cell. 1994b;77:817–827. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous