

Sequence-specific recognition of a subgenomic RNA promoter by a viral RNA polymerase
- PMID: 9326593
- PMCID: PMC23427
- DOI: 10.1073/pnas.94.21.11238
Sequence-specific recognition of a subgenomic RNA promoter by a viral RNA polymerase
Abstract
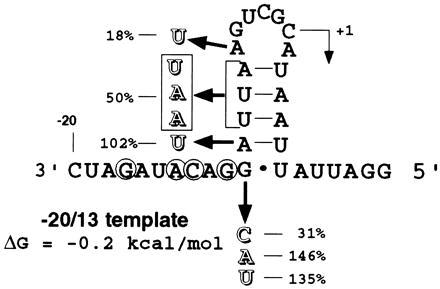
RNA templates of 33 nucleotides containing the brome mosaic virus (BMV) core subgenomic promoter were used to determine the promoter elements recognized by the BMV RNA-dependent RNA polymerase (RdRp) to initiate RNA synthesis. Nucleotides at positions -17, -14, -13, and -11 relative to the subgenomic initiation site must be maintained for interaction with the RdRp. Changes to every other nucleotide at these four positions allow predictions for the base-specific functional groups required for RdRp recognition. RdRp contact of the nucleotide at position -17 was suggested with a template competition assay. Comparison of the BMV subgenomic promoter to those from other plant and animal alphaviruses shows a remarkable degree of conservation of the nucleotides required for BMV subgenomic RNA synthesis. We show that the RdRp of the plant-infecting BMV is capable of accurately, albeit inefficiently, initiating RNA synthesis from the subgenomic promoter of the animal-infecting Semliki Forest virus. The sequence-specific recognition of RNA by the BMV RdRp is analogous to the recognition of DNA promoters by DNA-dependent RNA polymerases.
Figures
References
-
- Goldbach R, LeGall O, Welink J. Semin Virol. 1991;2:19–25.
-
- Miller W A, Dreher T W, Hall T C. Nature (London) 1985;313:68–70. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
