

Molecular immune responses of the mosquito Anopheles gambiae to bacteria and malaria parasites
- PMID: 9326640
- PMCID: PMC23521
- DOI: 10.1073/pnas.94.21.11508
Molecular immune responses of the mosquito Anopheles gambiae to bacteria and malaria parasites
Abstract
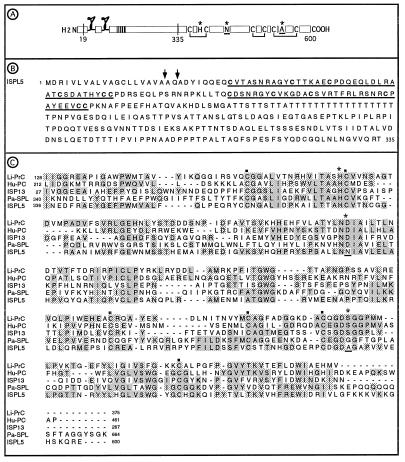
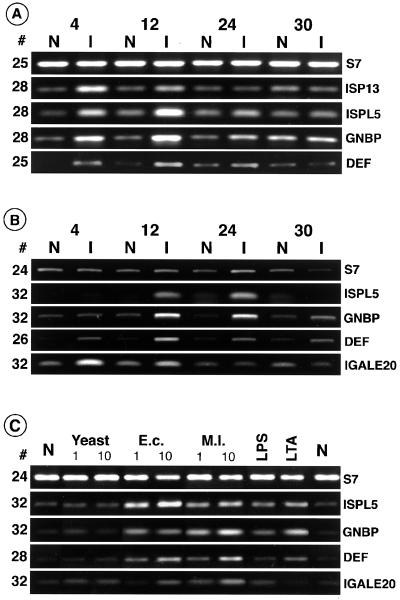
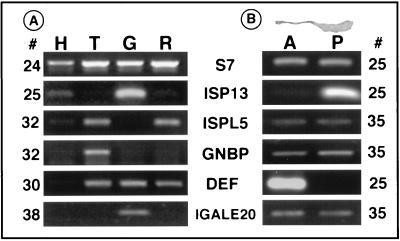
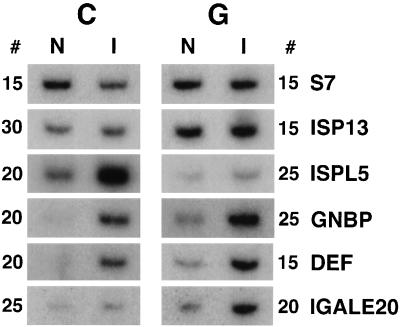
Immune responses of the malaria vector mosquito Anopheles gambiae were monitored systematically by the induced expression of five RNA markers after infection challenge. One newly isolated marker encodes a homologue of the moth Gram-negative bacteria-binding protein (GNBP), and another corresponds to a serine protease-like molecule. Additional previously described markers that respond to immune challenge encode the antimicrobial peptide defensin, a putative galactose lectin, and a putative serine protease. Specificity of the immune responses was indicated by differing temporal patterns of induction of specific markers in bacteria-challenged larvae and adults, and by variations in the effectiveness of different microorganisms and their components for marker induction in an immune-responsive cell line. The markers exhibit spatially distinct patterns of expression in the adult female mosquito. Two of them are highly expressed in different regions of the midgut, one in the anterior and the other in the posterior midgut. Marker induction indicates a significant role of the midgut in insect innate immunity. Immune responses to the penetration of the midgut epithelium by a malaria parasite occur both within the midgut itself and elsewhere in the body, suggesting an immune-related signaling process.
Figures

Comment in
-
Immune responsiveness in vector insects.Proc Natl Acad Sci U S A. 1997 Oct 14;94(21):11152-3. doi: 10.1073/pnas.94.21.11152. Proc Natl Acad Sci U S A. 1997. PMID: 9326575 Free PMC article. Review. No abstract available.
References
-
- Hoffman J A, Reichhard J-M, Hetru C. Curr Opin Immunol. 1996;8:8–13. - PubMed
-
- Hultmark D. Trends Genet. 1993;9:178–183. - PubMed
-
- Cociancich S, Ghazi A, Hetru C, Hoffman J, A, Letellier L. J Biol Chem. 1993;268:19239–19245. - PubMed
-
- Paskewitz S M, Christensen B M. In: The Biology of Disease Vectors. Beaty B J, Marquardt W C, editors; Beaty B J, Marquardt W C, editors. Boulder: Univer. Press of Colorado; 1997. pp. 371–392.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
