

The PAG gene product, a stress-induced protein with antioxidant properties, is an Abl SH3-binding protein and a physiological inhibitor of c-Abl tyrosine kinase activity
- PMID: 9334312
- PMCID: PMC316562
- DOI: 10.1101/gad.11.19.2456
The PAG gene product, a stress-induced protein with antioxidant properties, is an Abl SH3-binding protein and a physiological inhibitor of c-Abl tyrosine kinase activity
Abstract
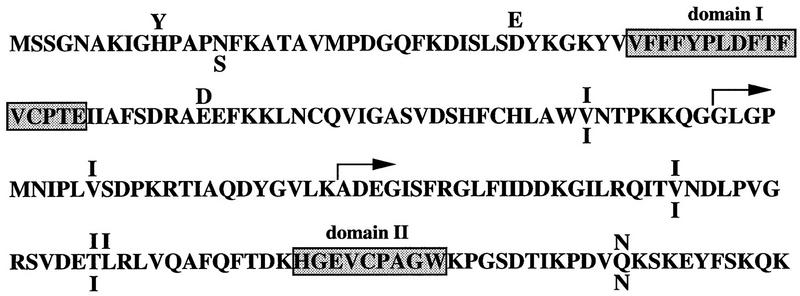
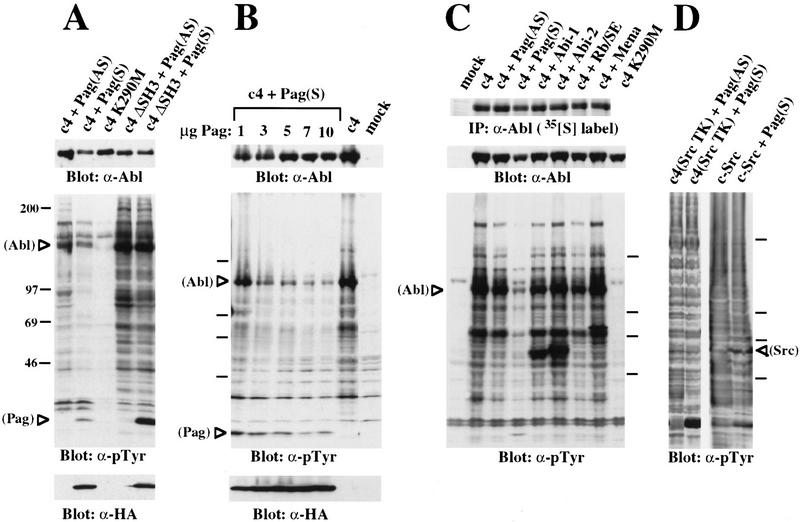
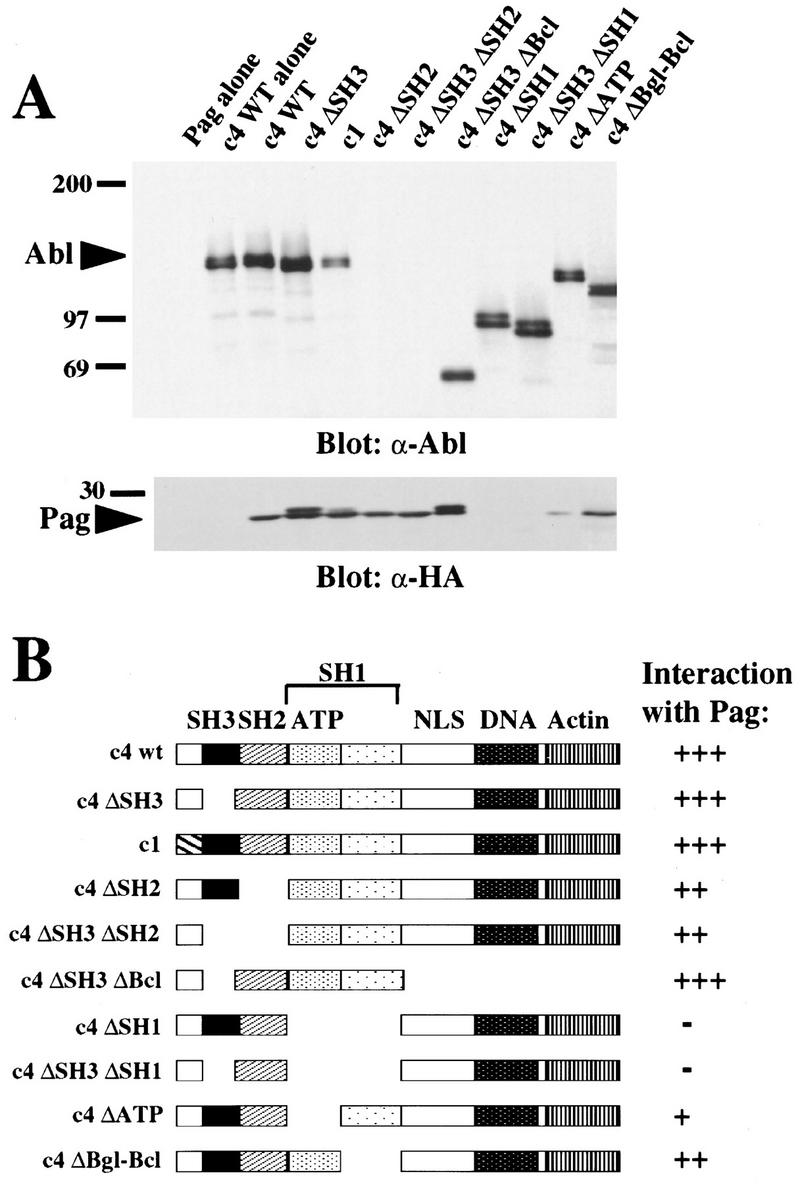
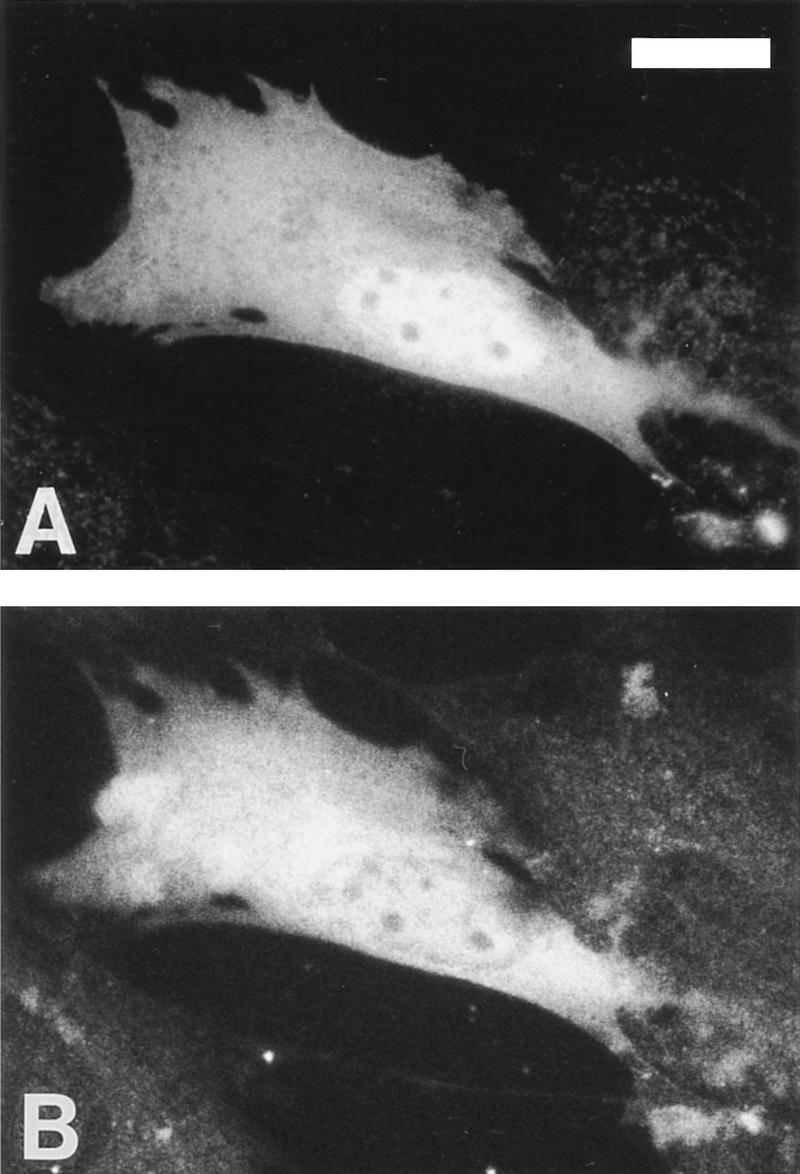
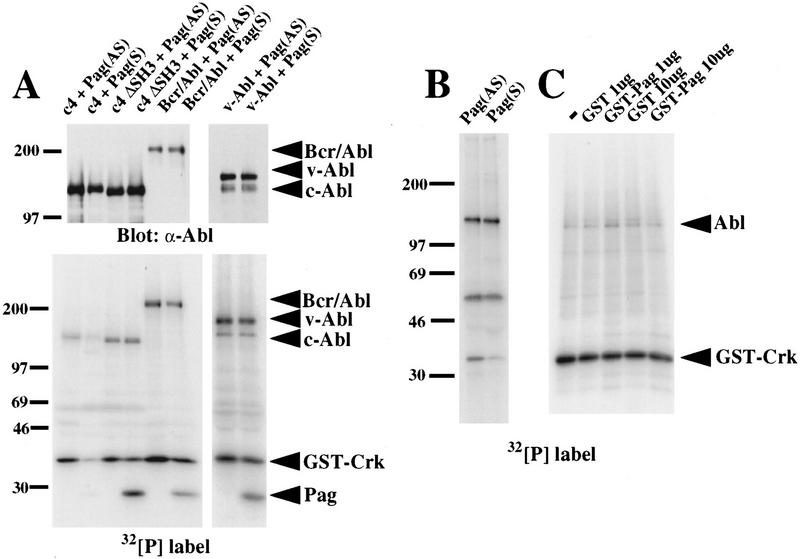
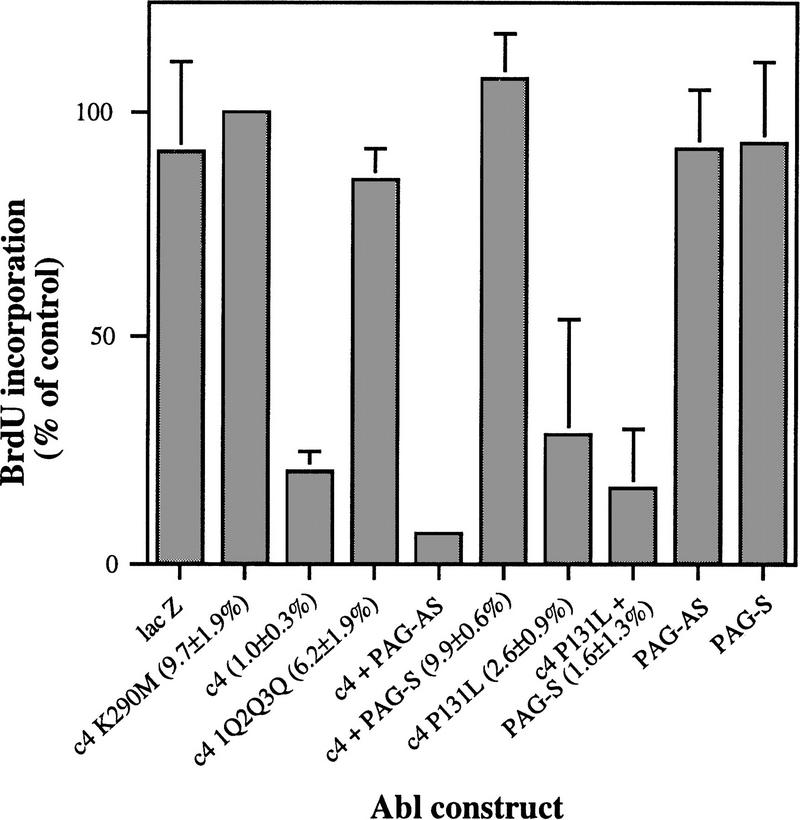
Biochemical and genetic evidence suggests that the tyrosine kinase activity of c-Abl is tightly regulated in vivo by a cellular factor binding to the Src homology 3 (SH3) domain of Abl. We used the yeast two-hybrid system to identify a gene, PAG, whose protein product (Pag) interacts specifically with the Abl SH3 domain. Pag, also known as macrophage 23-kD stress protein (MSP23), is a member of a novel family of proteins with antioxidant activity implicated in the cellular response to oxidative stress and in control of cell proliferation and differentiation. In a co-expression assay, Pag associates with c-Abl in vivo and inhibits tyrosine phosphorylation induced by overexpression of c-Abl. Inhibition requires the Abl SH3 and kinase domains and is not observed with other Abl SH3-binding proteins. Expression of Pag also inhibits the in vitro kinase activity of c-Abl, but not SH3-mutated Abl or v-Abl. When transfected in NIH-3T3 cells, Pag is localized to nucleus and cytoplasm and rescues the cytostatic effect induced by c-Abl. These observations suggest Pag is a physiological inhibitor of c-Abl in vivo.
Figures

References
-
- Adzhubei AA, Sternberg MJ. Left-handed poly-proline II helices commonly occur in globular proteins. J Mol Biol. 1993;229:472–493. - PubMed
-
- Andreotti AH, Bunnell SC, Feng S, Berg LJ, Schreiber SL. Regulatory intramolecular association in a tyrosine kinase of the Tec family. Nature. 1997;385:93–97. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous