

Transcription elongation factor P-TEFb is required for HIV-1 tat transactivation in vitro
- PMID: 9334325
- PMCID: PMC316609
- DOI: 10.1101/gad.11.20.2622
Transcription elongation factor P-TEFb is required for HIV-1 tat transactivation in vitro
Abstract
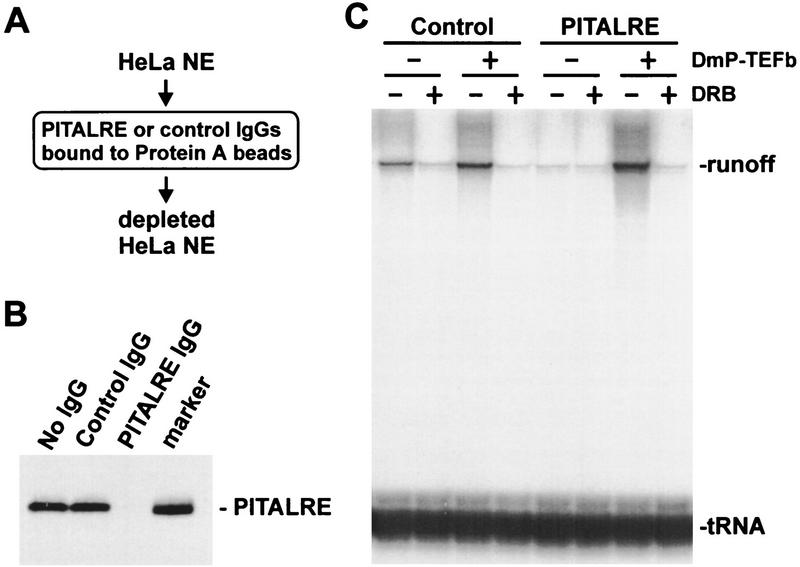
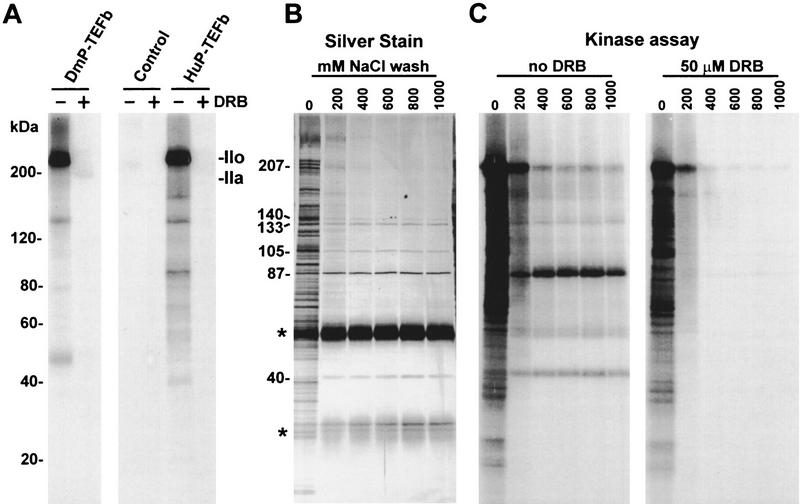
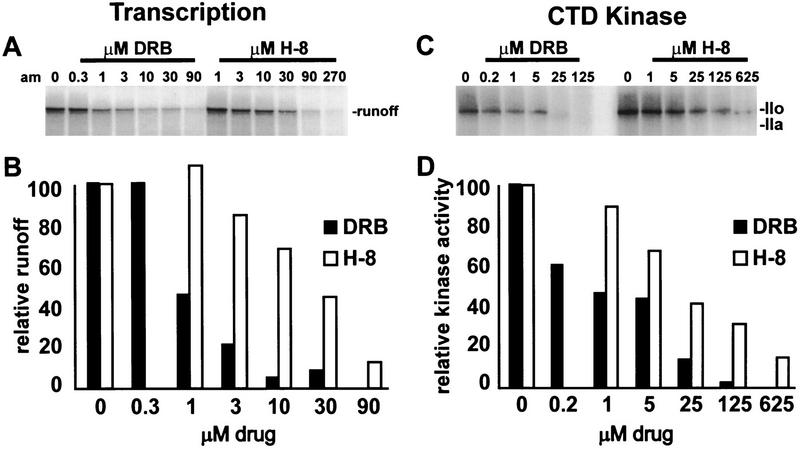
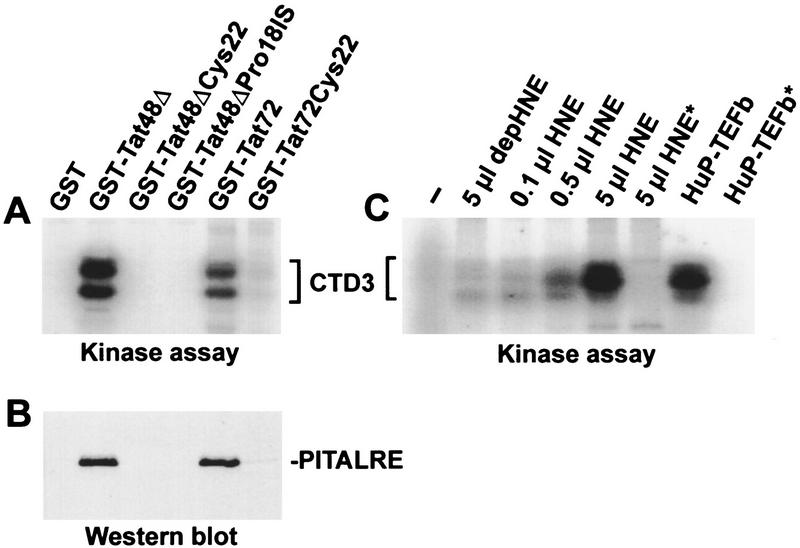
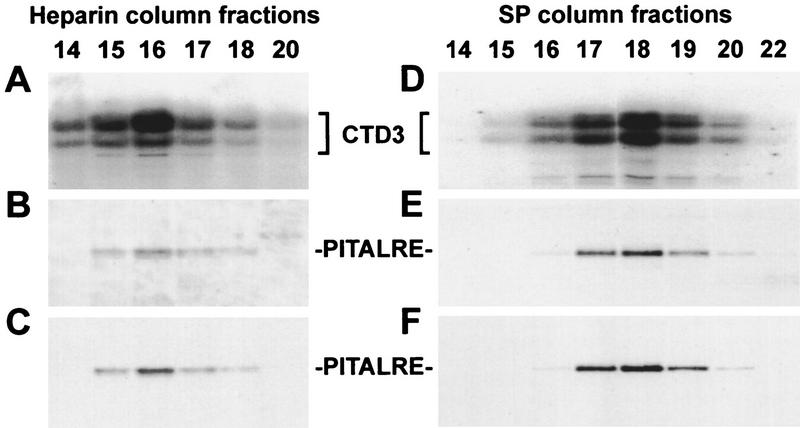
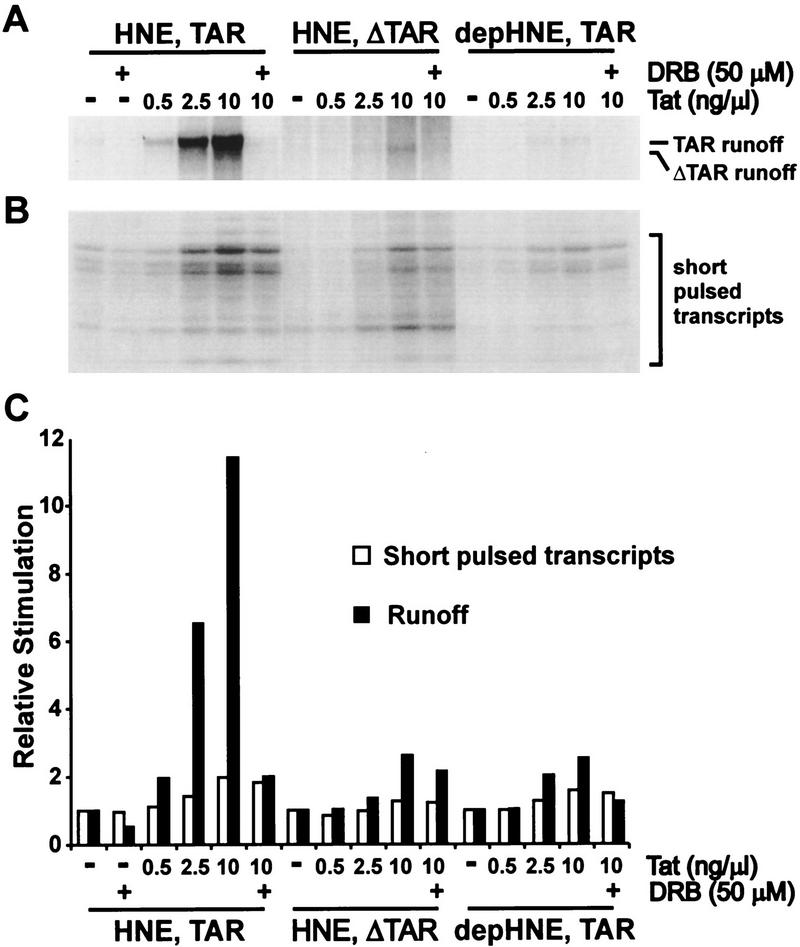
P-TEFb is a key regulator of the process controlling the processivity of RNA polymerase II and possesses a kinase activity that can phosphorylate the carboxy-terminal domain of the largest subunit of RNA polymerase II. Here we report the cloning of the small subunit of Drosophila P-TEFb and the finding that it encodes a Cdc2-related protein kinase. Sequence comparison suggests that a protein with 72% identity, PITALRE, could be the human homolog of the Drosophila protein. Functional homology was suggested by transcriptional analysis of an RNA polymerase II promoter with HeLa nuclear extract depleted of PITALRE. Because the depleted extract lost the ability to produce long DRB-sensitive transcripts and this loss was reversed by the addition of purified Drosophila P-TEFb, we propose that PITALRE is a component of human P-TEFb. In addition, we found that PITALRE associated with the activation domain of HIV-1 Tat, indicating that P-TEFb is a Tat-associated kinase (TAK). An in vitro transcription assay demonstrates that the effect of Tat on transcription elongation requires P-TEFb and suggests that the enhancement of transcriptional processivity by Tat is attributable to enhanced function of P-TEFb on the HIV-1 LTR.
Figures

References
-
- Aso T, Lane WS, Conaway JW, Conaway RC. Elongin (SIII): A multisubunit regulator of elongation by RNA polymerase II. Science. 1995;269:1439–1443. - PubMed
-
- Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K. Current protocols in molecular biology. New York, NY: Greene Publishing Associates/Wiley-Intersience; 1989.
-
- Bentley DL. Regulation of transcriptional elongation by RNA polymerase II. Curr Opin Genet Dev. 1995;5:210–216. - PubMed
-
- Braddock M, Thorburn AM, Kingsman AJ, Kingsman SM. Blocking of Tat-dependent HIV-1 RNA modification by an inhibitor of RNA polymerase II processivity. Nature. 1991;350:439–441. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous