

doi: 10.1083/jcb.139.2.459.
The yeast motor protein, Kar3p, is essential for meiosis I
Affiliations
- PMID: 9334348
- PMCID: PMC2139793
- DOI: 10.1083/jcb.139.2.459
Item in Clipboard
The yeast motor protein, Kar3p, is essential for meiosis I
J Cell Biol.
.
Abstract
The recognition and alignment of homologous chromosomes early in meiosis is essential for their subsequent segregation at anaphase I; however, the mechanism by which this occurs is unknown. We demonstrate here that, in the absence of the molecular motor, Kar3p, meiotic cells are blocked with prophase monopolar microtubule arrays and incomplete synaptonemal complex (SC) formation. kar3 mutants exhibit very low levels of heteroallelic recombination. kar3 mutants do produce double-strand breaks that act as initiation sites for meiotic recombination in yeast, but at levels severalfold reduced from wild-type. These data are consistent with a meiotic role for Kar3p in the events that culminate in synapsis of homologues.
Figures

Analysis of tubulin and DNA staining pattern of KAR3 and kar3 cells throughout meiosis. Aliquots of isogenic wild-type (DJ1 + pD154.12) and kar3 (DJ1) cells were fixed at the indicated times after transfer to sporulation medium and stained with antitubulin antibodies and the DNA-specific dye DAPI. Antitubulin and DAPI images of representative cells are shown for both wild-type (top) and kar3 (bottom) time courses. Arrows in the 5-h kar3 samples indicate pronounced cytoplasmic projections. The arrow in the 24-h kar3 panel identifies a highly abnormal spindle. Small numbers of unusual spindles were observed in all time points for the kar3 culture. Bar, 10 μm.

kar3 mutant cells are blocked before metaphase I. The frequency of cells exhibiting unquestionable postprophase I morphology was determined for both the wild-type and kar3 cells. n ⩾ 28 for each time point.
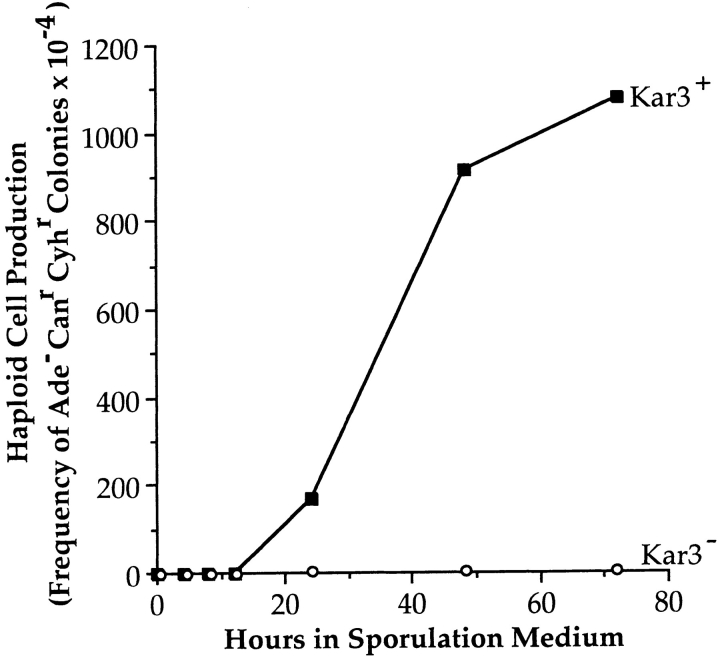
kar3 mutants fail to produce haploid meiotic products. Isogenic kar3 (TC12) and KAR3 (DJ1 + pD154.12) cultures heterozygous for ade2, cyh2, and can1 were pregrown and induced to undergo meiosis as described in Materials and Methods. At the times indicated, aliquots were removed and plated onto YPD and CM-Arg + cyclohexamide + canavanine. The parameter plotted is the fraction of Ade−, Canr, Cyhr CFUs at each time point relative to the total CFUs on YPD plates. For the kar3 mutants, at least 13,000 cells were plated at each time point. Ade−, Cyhr, Canr colonies were never observed for the kar3 mutants.
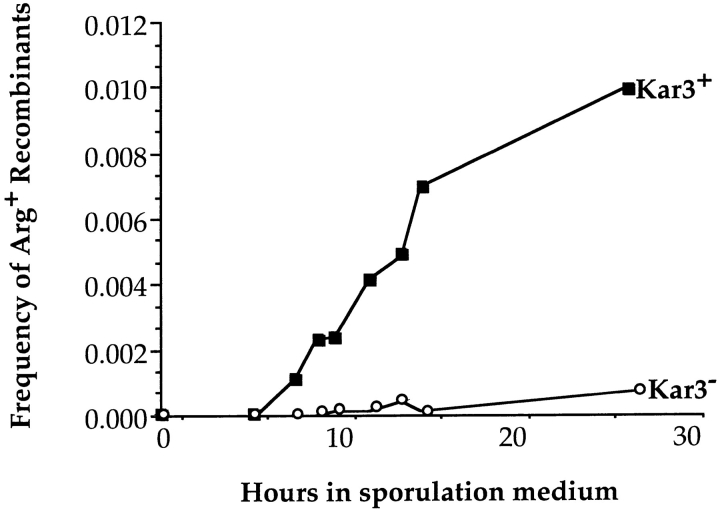
kar3 cells undergo interhomologue recombination at severely reduced levels. Isogenic cultures of KAR3 (Dd519 + MR820) and kar3 (Dd519) strains, heterozygous for a pair of arg4 heteroalleles (arg4RV and arg4Δ42), were induced to undergo synchronous meiosis as described in Materials and Methods. Aliquots were removed at the indicated times and plated onto CM-Arg to determine the number of Arg+ recombinants. Aliquots were also plated onto YPD or CM to determine total CFUs. Plotted is the ratio of Arg+ CFUs to total CFUs. There was little loss of viability throughout the course of this experiment. The kar3 cells showed ∼5% of wild-type levels of recombination at 27 h.
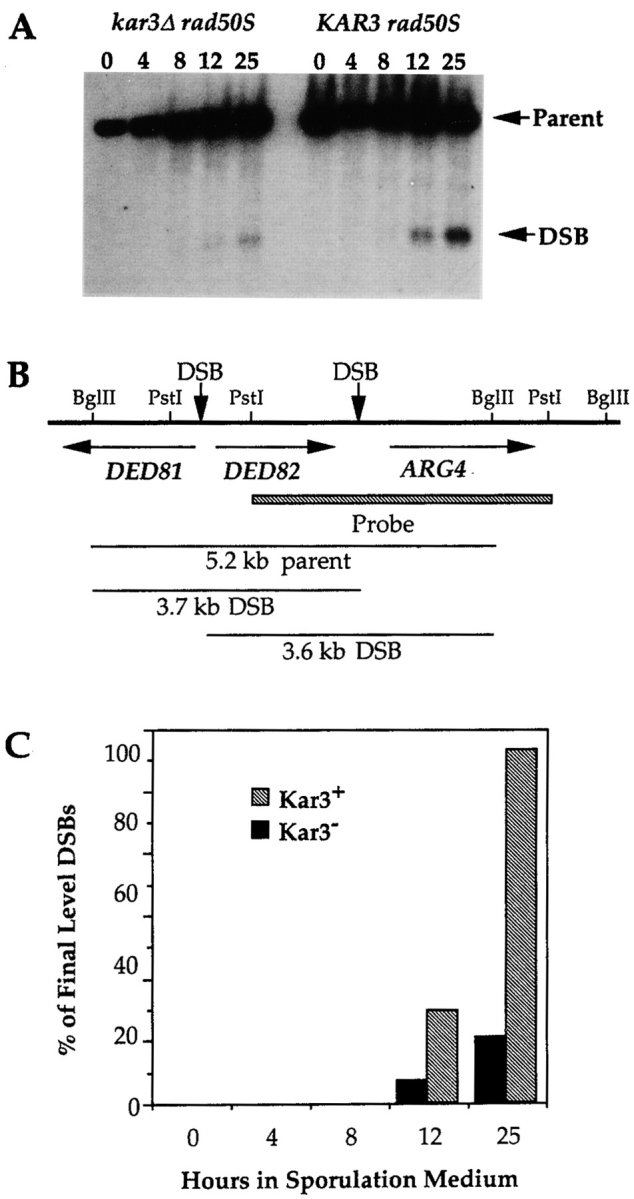
kar3 cells are capable of initiating DSBs. Isogenic KAR3 (DC99-1) or kar3 (DC99-1C) cultures, homozygous for the rad50S-K181 allele were pregrown and induced to undergo synchronous meiosis. Aliquots were removed at the number of hours indicated after transfer to sporulation medium and DNA was extracted. The DNA was digested with BglII, subjected to electrophoresis on a 0.7% agarose gel, and analyzed by Southern blot hybridization using as a probe the ARG4 sequences indicated in (B). (A) The predominant DSB and parental fragments are labeled with arrows. (C) Quantification of the DSB products. The fraction of radioactivity in each lane that was localized to the DSB products was quantified. The percent of DSB product in each lane was normalized to the final level of DSB product produced in the KAR3 strain. In the kar3 mutants, the DSBs accumulate to 18.6% of levels detected in the isogenic KAR3 strain.

Zip1 staining pattern of kar3 cells. Aliquots of isogenic KAR3 (Dd519 + MR820) and kar3 (Dd519) meiotic cultures were harvested at pachytene, 14 h after induction of sporulation. Nuclei were spread and stained with anti-Zip1 and the DNA-specific dye, DAPI. (a) The left panel shows the Zip1 staining pattern of a typical wild-type pachytene nucleus; the corresponding staining with DAPI is shown on the right. (b and c) The left-hand panels are Zip1-stained nuclei representative of those with dots or elongated signals. The corresponding DAPI images are shown on the right. Brightly stained foci indicative of polycomplexes were observed in 25% of the kar3 cells, and 2.5% of the wild-type cells (arrows). Bar, 1 μm.

Electron micrographs of pachytene nuclei. Isogenic KAR3 (Dd519 + MR820) and kar3 (Dd519) nuclei were harvested at pachytene and spread as indicated in Materials and Methods. (A and B) SCs from wild-type nuclei. (C–F) Examples of representative SC morphologies detected in kar3 cells. Some cells exhibit a large degree of synapsis (C), yet others show little to no SC (E). Duplicated, but unseparated SPBs from kar3 cells are apparent in E and F (arrows). Bar, 1 μm.
References
-
- Afshar K, Barton NR, Hawley RS, Goldstein LSB. DNA binding and meiotic chromosomal localization of the Drosophila nod kinesin-like protein. Cell. 1995;81:129–138. - PubMed
-
- Alani E, Padmore R, Kleckner N. Analysis of wild-type and rad50mutants of yeast suggests an intimate relationship between meiotic chromosome synapsis and recombination. Cell. 1990;61:419–436. - PubMed
-
- Bascom-Slack, C.A., L.O. Ross, and D.S. Dawson. 1997. Chiasmata, crossovers, and meiotic chromosome segregation. In Advances in Genetics. Vol. 35. J. Hall and J. Dunlap, editors. Academic Press, Inc., San Diego, CA. 253–284. - PubMed
-
- Bishop DK, Park D, Xu L, Kleckner N. DMC1: a meiosis-specific yeast homolog of E. coli recArequired for recombination, synaptonemal complex formation, and cell cycle progression. Cell. 1992;69:439–456. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
