

Laminin alpha1 chain synthesis in the mouse developing lung: requirement for epithelial-mesenchymal contact and possible role in bronchial smooth muscle development
- PMID: 9334356
- PMCID: PMC2139794
- DOI: 10.1083/jcb.139.2.553
Laminin alpha1 chain synthesis in the mouse developing lung: requirement for epithelial-mesenchymal contact and possible role in bronchial smooth muscle development
Abstract
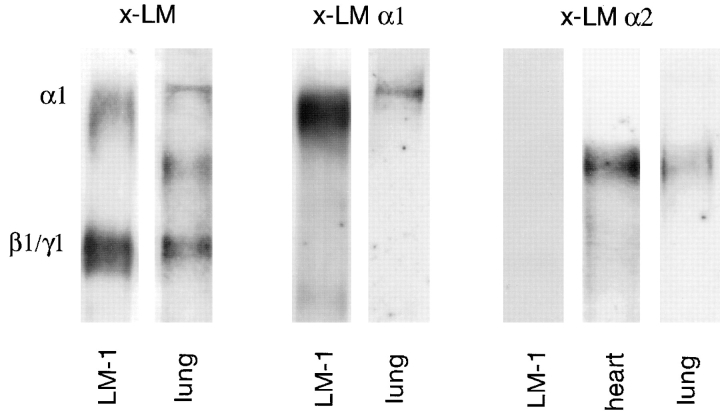
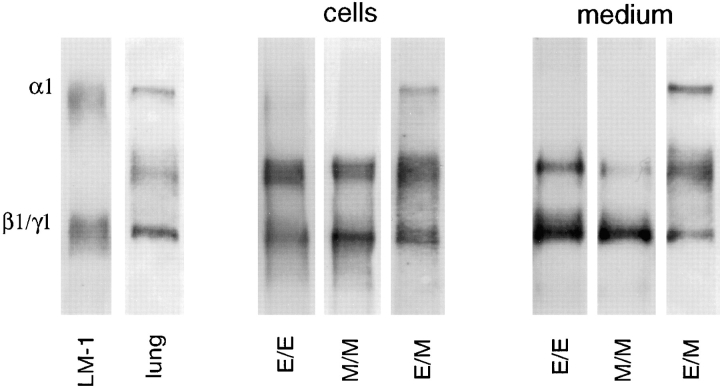
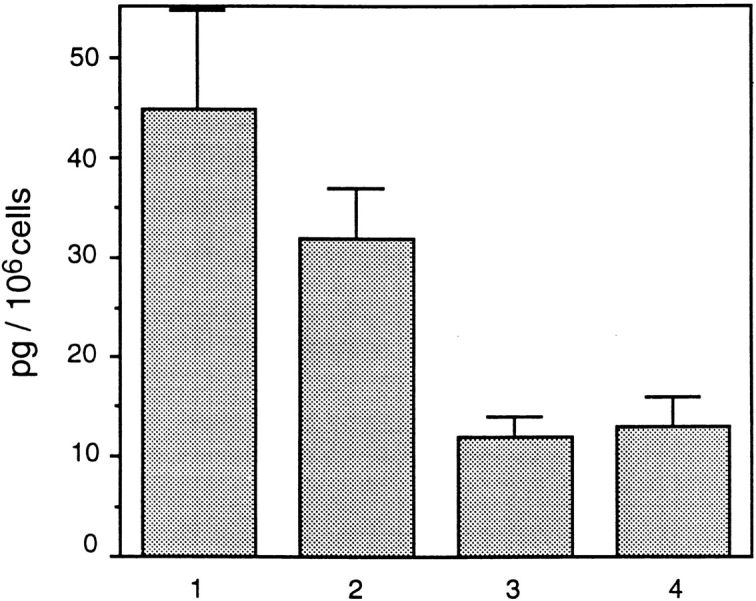
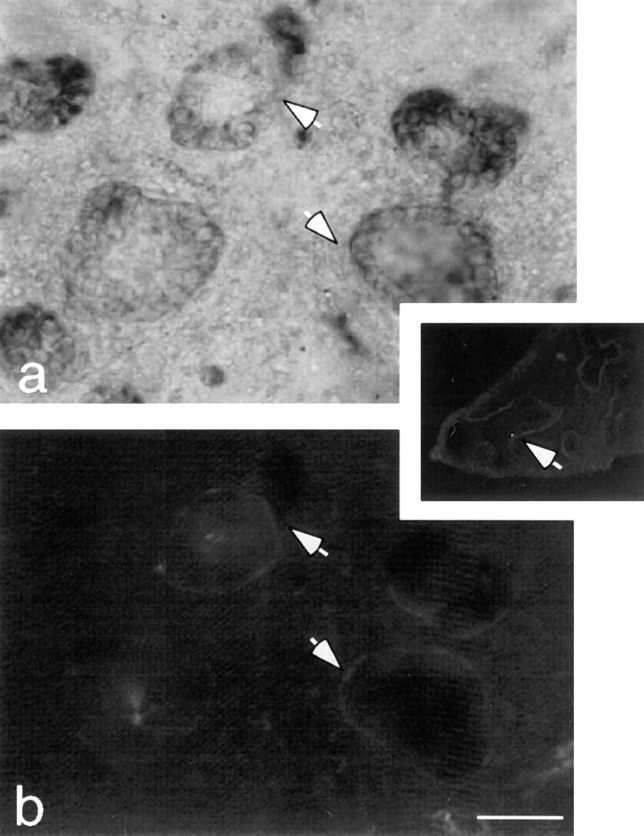
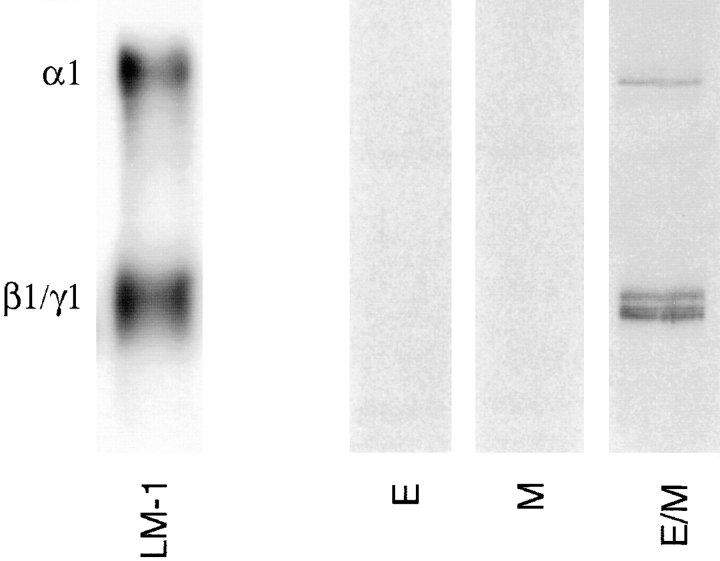
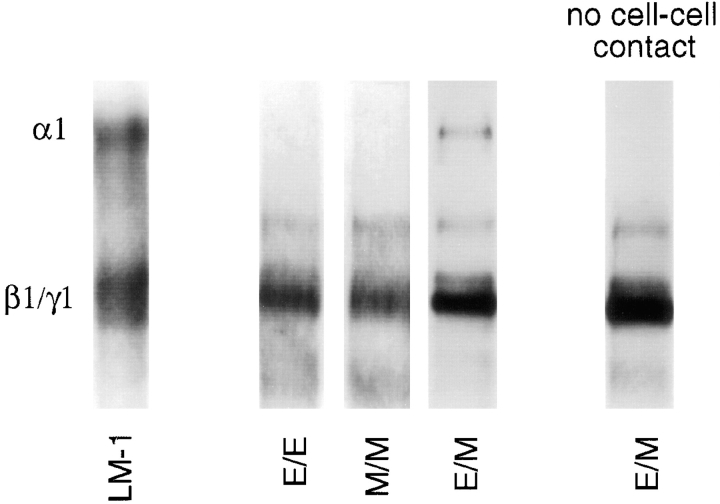
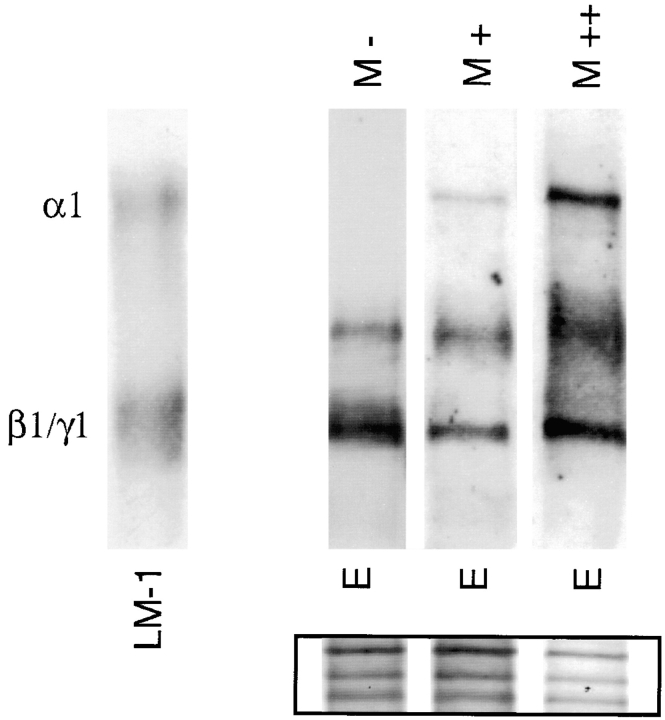
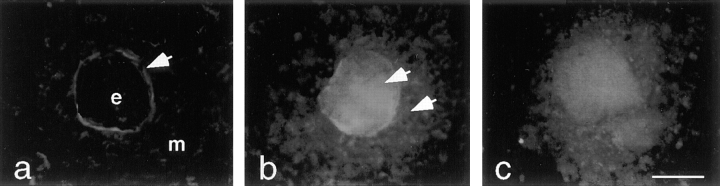
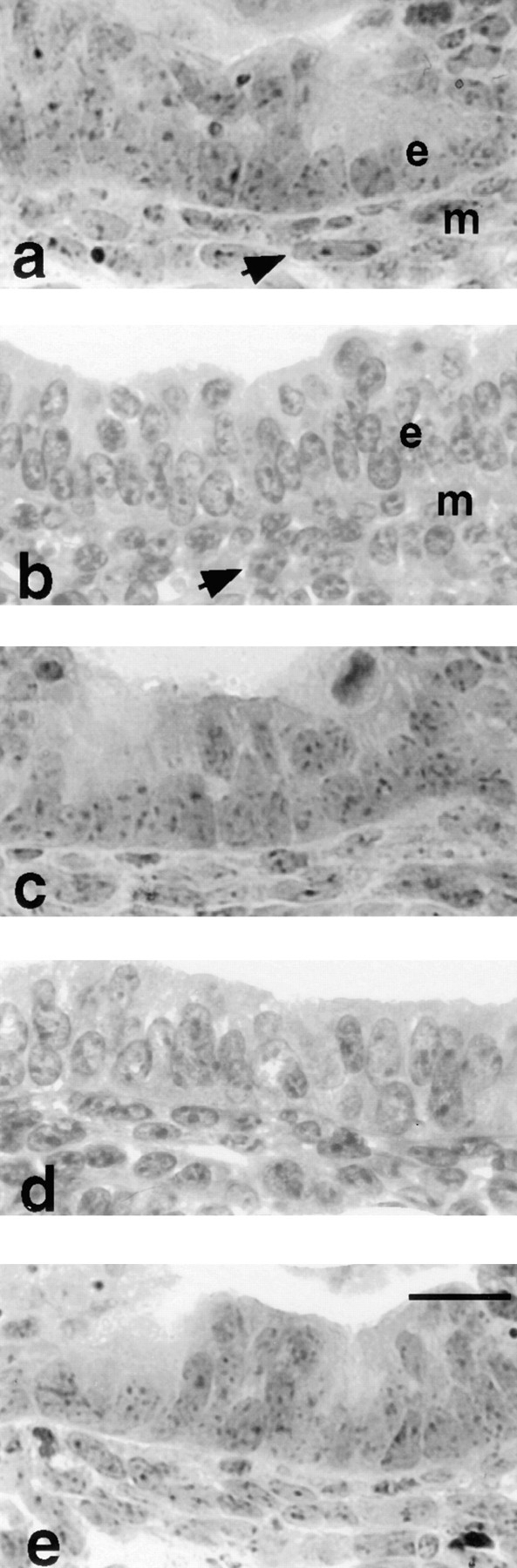
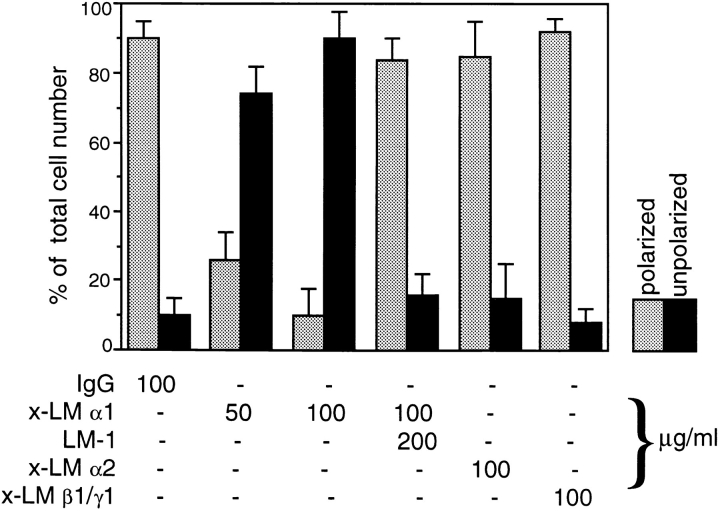
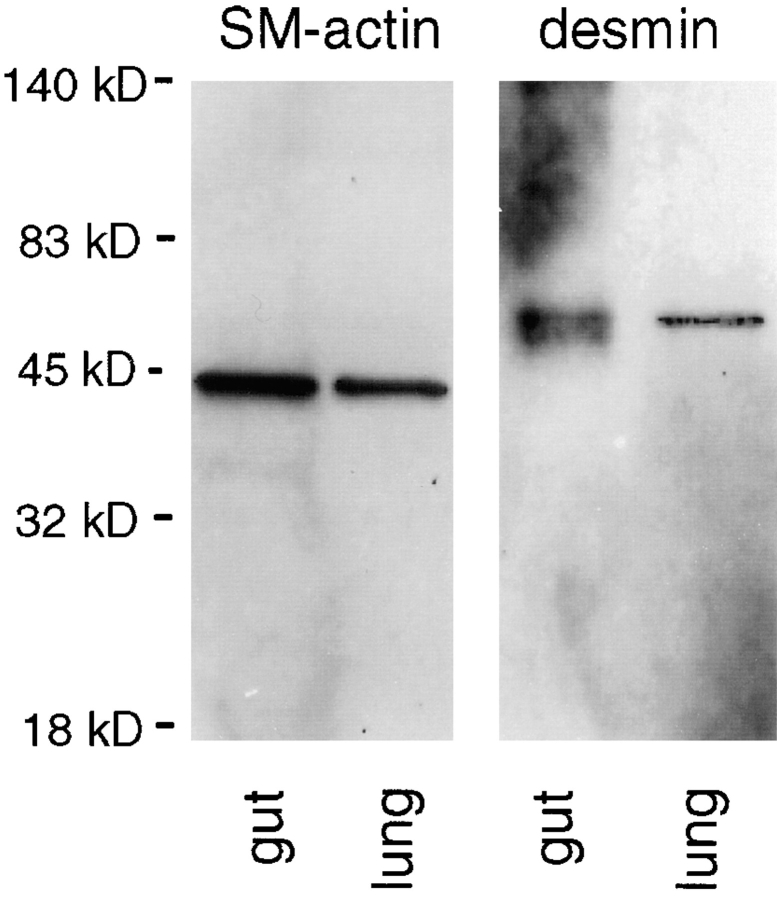
Laminins, the main components of basement membranes, are heterotrimers consisting of alpha, beta, and gamma polypeptide chains linked together by disulfide bonds. Laminins-1 and -2 are both composed of beta1 and gamma1 chains and differ from each other on their alpha chain, which is alpha1 and alpha2 for laminin-1 and -2, respectively. The present study shows that whereas laminins-1 and -2 are synthesized in the mouse developing lung and in epithelial-mesenchymal cocultures derived from it, epithelial and mesenchymal monocultures lose their ability to synthesize the laminin alpha1 chain. Synthesis of laminin alpha1 chain however returns upon re-establishment of epithelial-mesenchymal contact. Cell-cell contact is critical, since laminin alpha1 chain is not detected in monocultures exposed to coculture-conditioned medium or in epithelial-mesenchymal cocultures in which heterotypic cell-cell contact is prevented by an interposing filter. Immunohistochemical studies on cocultures treated with brefeldin A, an inhibitor of protein secretion, indicated both epithelial and mesenchymal cells synthesize laminin alpha1 chain upon heterotypic cell- cell contact. In a set of functional studies, embryonic lung explants were cultured in the presence of monoclonal antibodies to laminin alpha1, alpha2, and beta/gamma chains. Lung explants exposed to monoclonal antibodies to laminin alpha1 chain exhibited alterations in peribronchial cell shape and decreased smooth muscle development, as indicated by low levels of smooth muscle alpha actin and desmin. Taken together, our studies suggest that laminin alpha1 chain synthesis is regulated by epithelial-mesenchymal interaction and may play a role in airway smooth muscle development.
Figures

References
-
- Boström H, Willetts K, Pekny M, Leveen P, Lindahl P, Hedstrand H, Pekna M, Hellström M, Gebre-Medhin S, Schalling M, et al. PDGF-A signaling is a critical event in lung alveolar myofibroblast development and alveogenesis. Cell. 1996;85:863–873. - PubMed
-
- Burgeson RE, Chiquet M, Deutzmann R, Ekblom P, Engel J, Kleinmann H, Martin GR, Meneguzzi G, Paulsson M, Sanes J, et al. A new nomenclature for the laminins. Matrix Biol. 1994;263:16536–16544. - PubMed
-
- Carlin BE, Durkin ME, Bender B, Jaffe R, Chung AE. Synthesis of laminin and entactin by F9 cells induced with retinoic acid and dibutyryl cyclic AMP. J Biol Chem. 1983;25:7729–7737. - PubMed
-
- Cunha GR, Battle E, Young P, Brody J, Donjacour A, Hayashi N, Kinbara H. Role of epithelial-mesenchymal interactions in the differentiation and spatial organization of visceral smooth muscle. Epithelial Cell Biol. 1992;2:76–83. - PubMed
