

Participation of the nuclear cap binding complex in pre-mRNA 3' processing
- PMID: 9342333
- PMCID: PMC23648
- DOI: 10.1073/pnas.94.22.11893
Participation of the nuclear cap binding complex in pre-mRNA 3' processing
Abstract
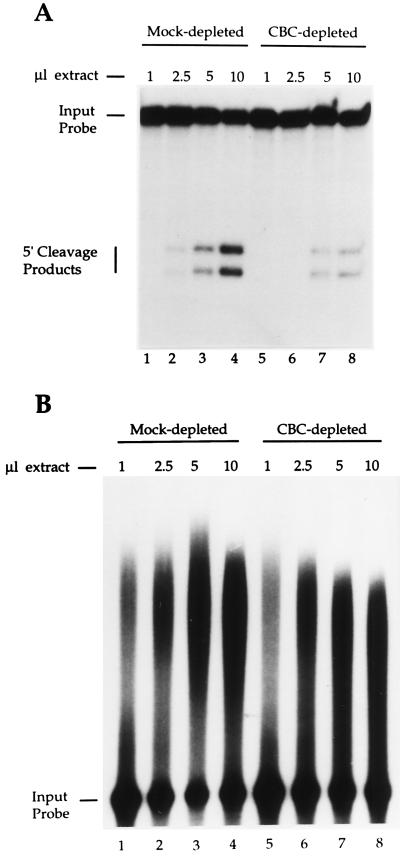
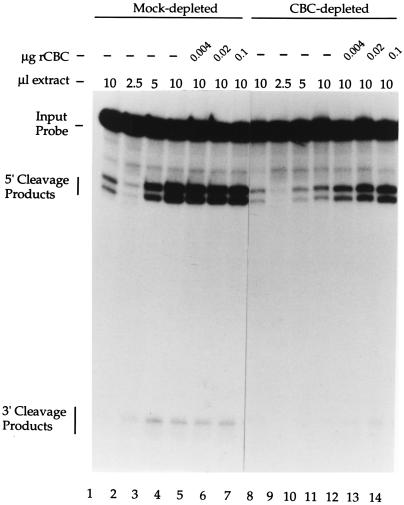
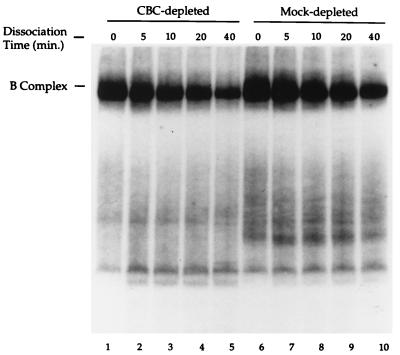
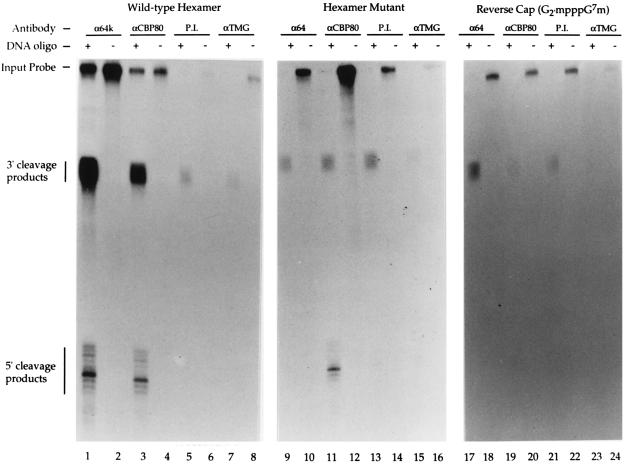
Communication between the 5' and 3' ends is a common feature of several aspects of eukaryotic mRNA metabolism. In the nucleus, the pre-mRNA 5' end is bound by the nuclear cap binding complex (CBC). This RNA-protein complex plays an active role in both splicing and RNA export. We provide evidence for participation of CBC in the processing of the 3' end of the message. Depletion of CBC from HeLa cell nuclear extract strongly reduced the endonucleolytic cleavage step of the cleavage and polyadenylation process. Cleavage was restored by addition of recombinant CBC. CBC depletion was found to reduce the stability of poly(A) site cleavage complexes formed in nuclear extract. We also provide evidence that the communication between the 5' and 3' ends of the pre-mRNA during processing is mediated by the physical association of the CBC/cap complex with 3' processing factors bound at the poly(A) site. These observations, along with previous data on the function of CBC in splicing, illustrate the key role played by CBC in pre-mRNA recognition and processing. The data provides further support for the hypothesis that pre-mRNAs and mRNAs may exist and be functional in the form of "closed-loops," due to interactions between factors bound at their 5' and 3' ends.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
