

Mapping and use of a sequence that targets DNA ligase I to sites of DNA replication in vivo
- PMID: 9348276
- PMCID: PMC2141708
- DOI: 10.1083/jcb.139.3.579
Mapping and use of a sequence that targets DNA ligase I to sites of DNA replication in vivo
Abstract
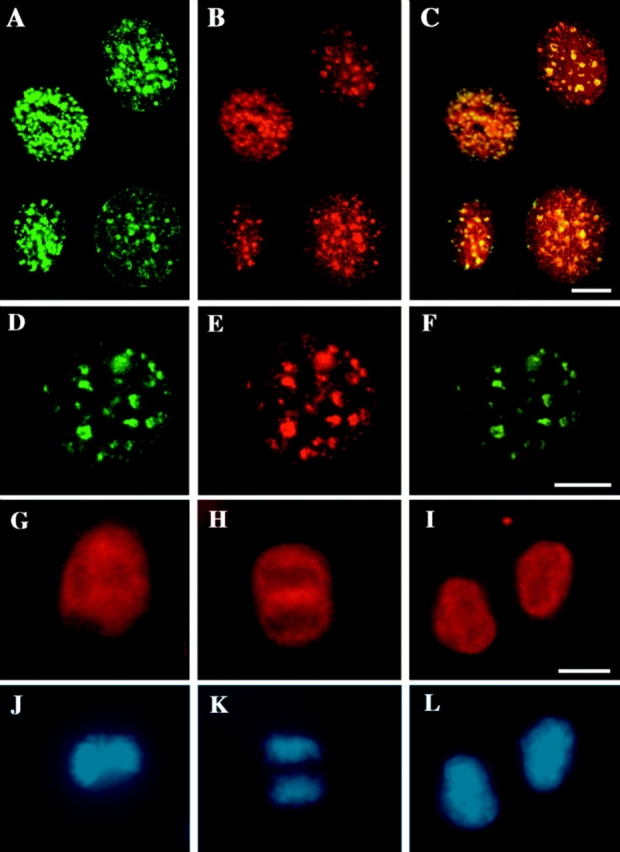
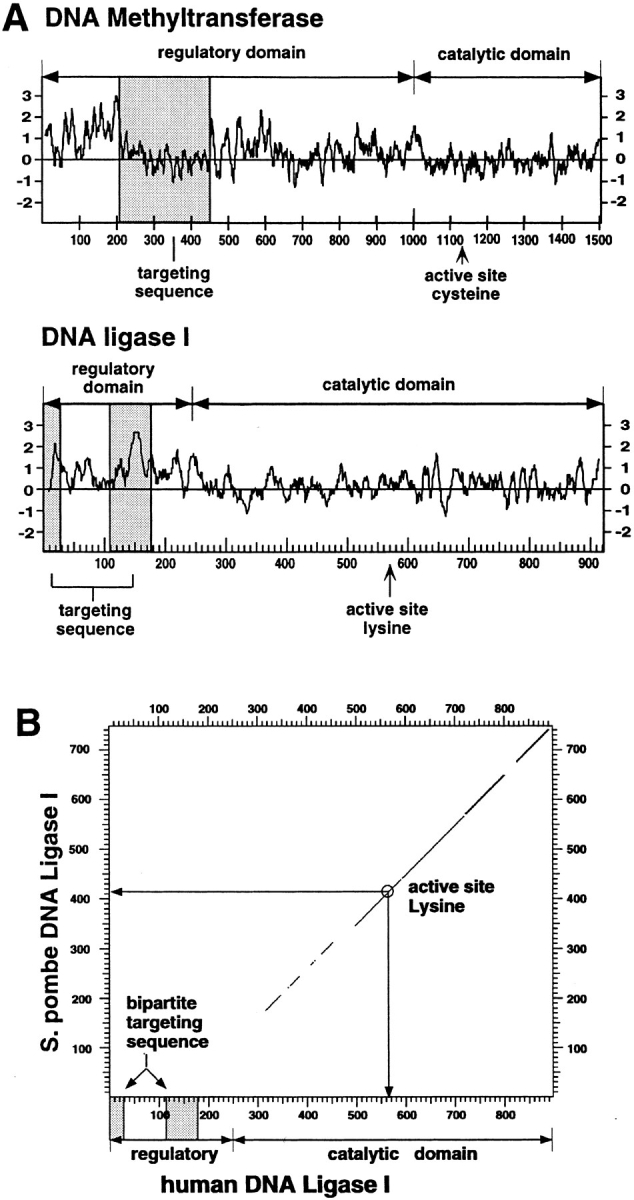
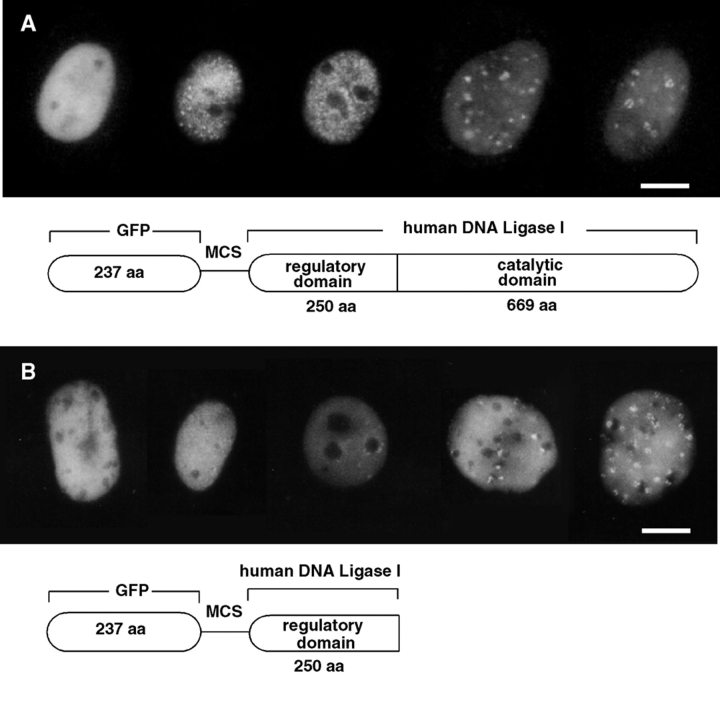
The mammalian nucleus is highly organized, and nuclear processes such as DNA replication occur in discrete nuclear foci, a phenomenon often termed "functional organization" of the nucleus. We describe the identification and characterization of a bipartite targeting sequence (amino acids 1-28 and 111-179) that is necessary and sufficient to direct DNA ligase I to nuclear replication foci during S phase. This targeting sequence is located within the regulatory, NH2-terminal domain of the protein and is dispensable for enzyme activity in vitro but is required in vivo. The targeting domain functions position independently at either the NH2 or the COOH termini of heterologous proteins. We used the targeting sequence of DNA ligase I to visualize replication foci in vivo. Chimeric proteins with DNA ligase I and the green fluorescent protein localized at replication foci in living mammalian cells and thus show that these subnuclear functional domains, previously observed in fixed cells, exist in vivo. The characteristic redistribution of these chimeric proteins makes them unique markers for cell cycle studies to directly monitor entry into S phase in living cells.
Figures




References
-
- Banks GR, Barker DG. DNA ligase-AMP adducts: identification of yeast DNA ligase polypeptides. Biochim Biophys Acta. 1985;826:180–185. - PubMed
-
- Barnes DE, Tomkinson AE, Lehmann AR, Webster AD, Lindahl T. Mutations in the DNA ligase I gene of an individual with immunodeficiencies and cellular hypersensitivity to DNA-damaging agents. Cell. 1992;69:495–503. - PubMed
-
- Bentley D, Selfridge J, Millar JK, Samuel K, Hole N, Ansell JD, Melton DW. DNA ligase I is required for fetal liver erythropoiesis but is not essential for mammalian cell viability. Nat Genet. 1996;13:489–491. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
