

ERM (ezrin/radixin/moesin)-based molecular mechanism of microvillar breakdown at an early stage of apoptosis
- PMID: 9348291
- PMCID: PMC2141718
- DOI: 10.1083/jcb.139.3.749
ERM (ezrin/radixin/moesin)-based molecular mechanism of microvillar breakdown at an early stage of apoptosis
Abstract
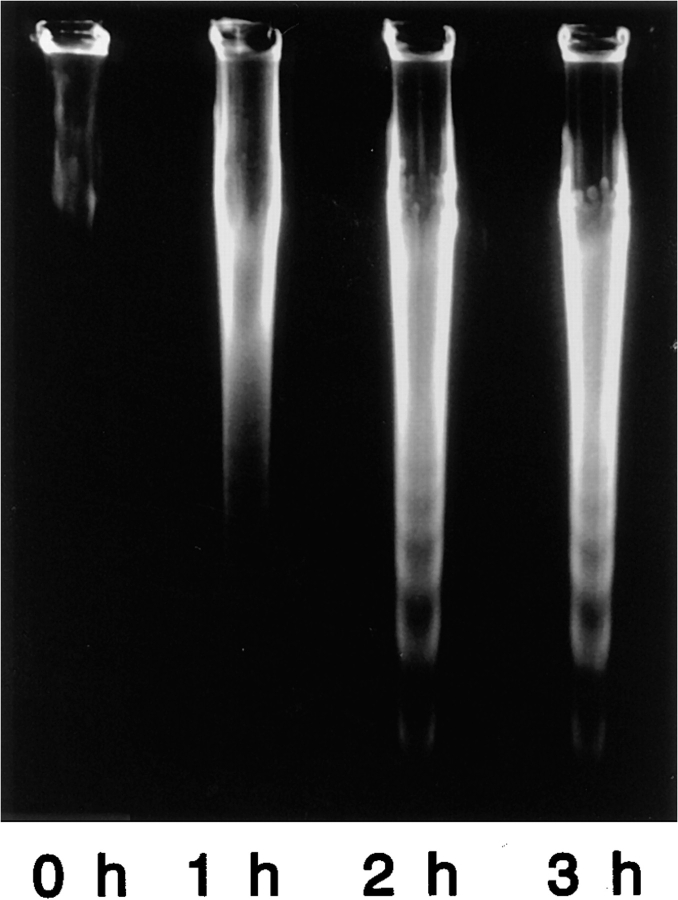
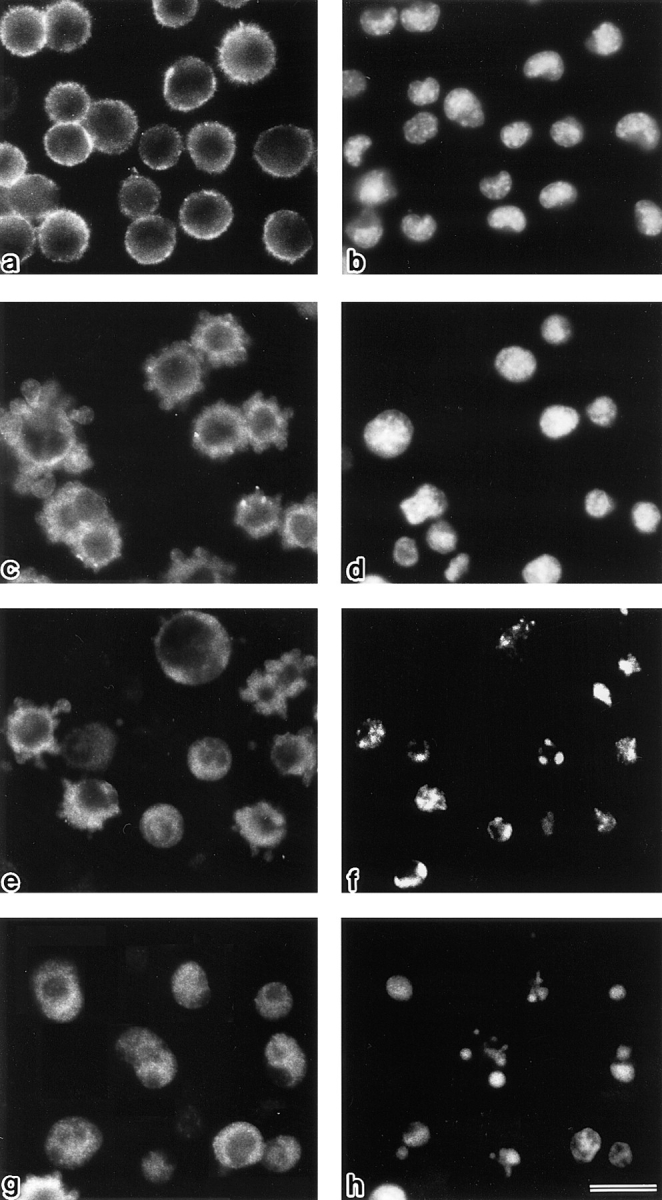
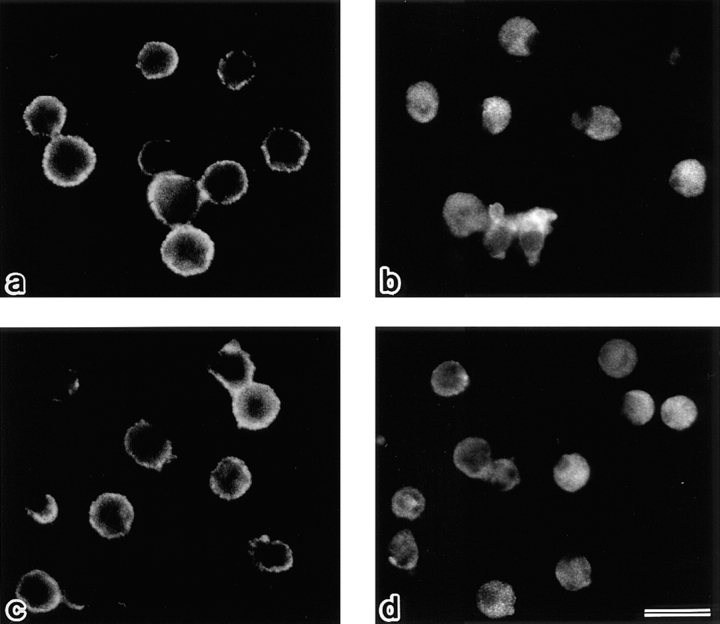
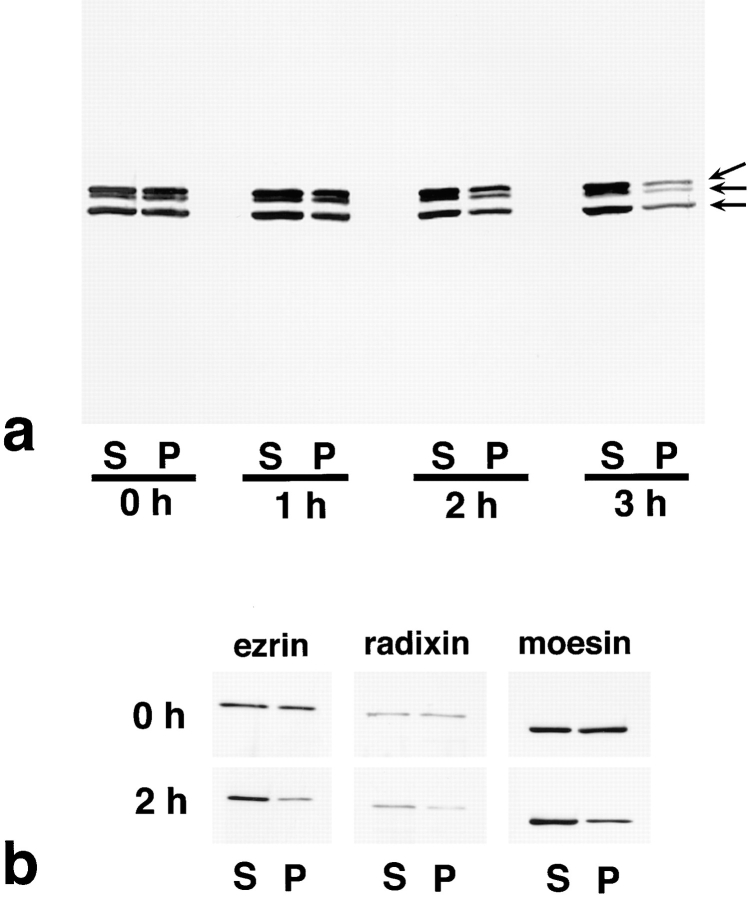
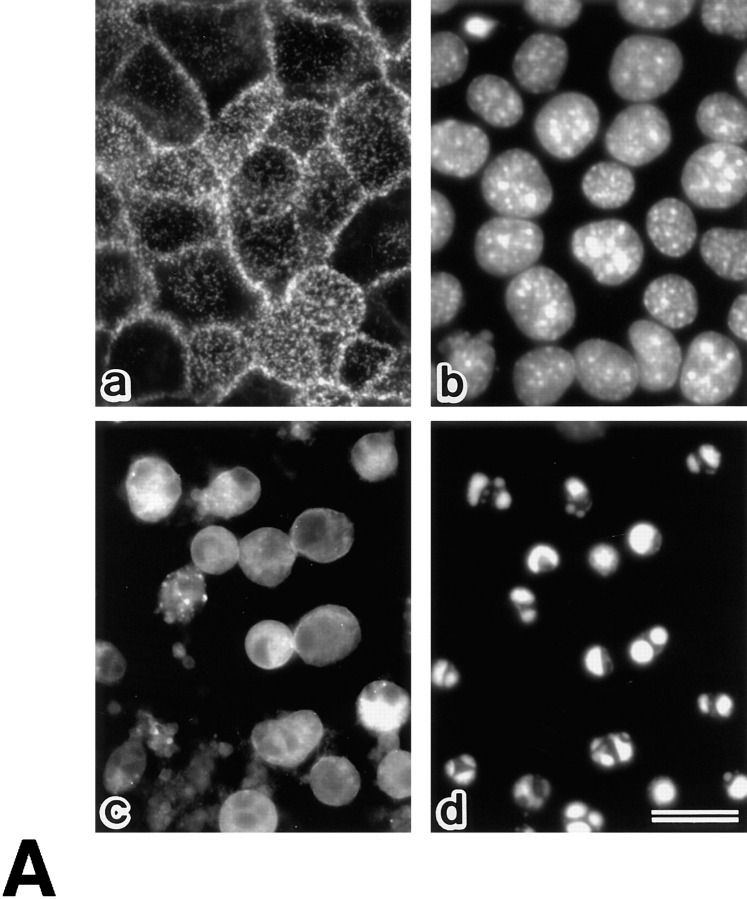
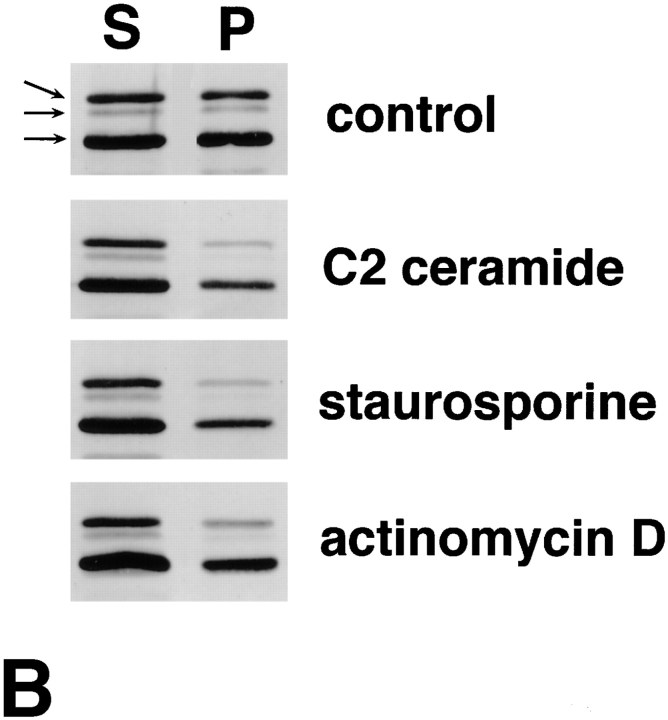
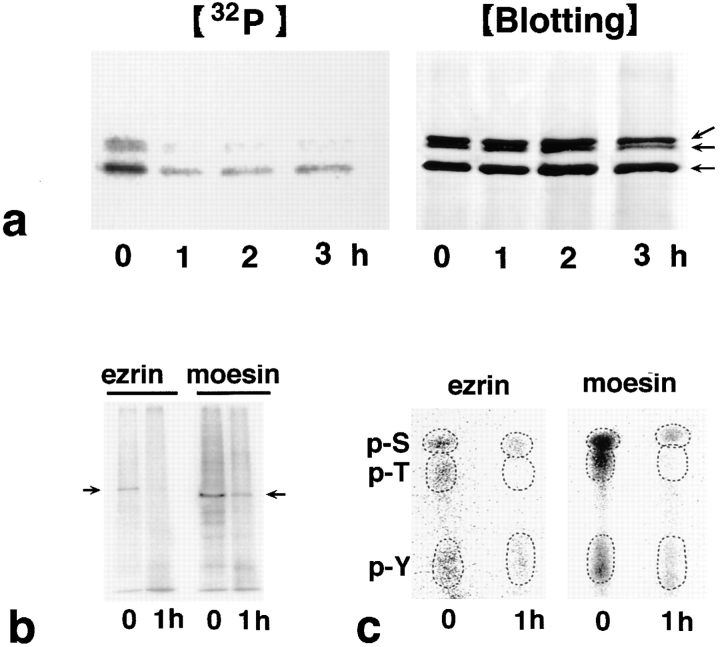
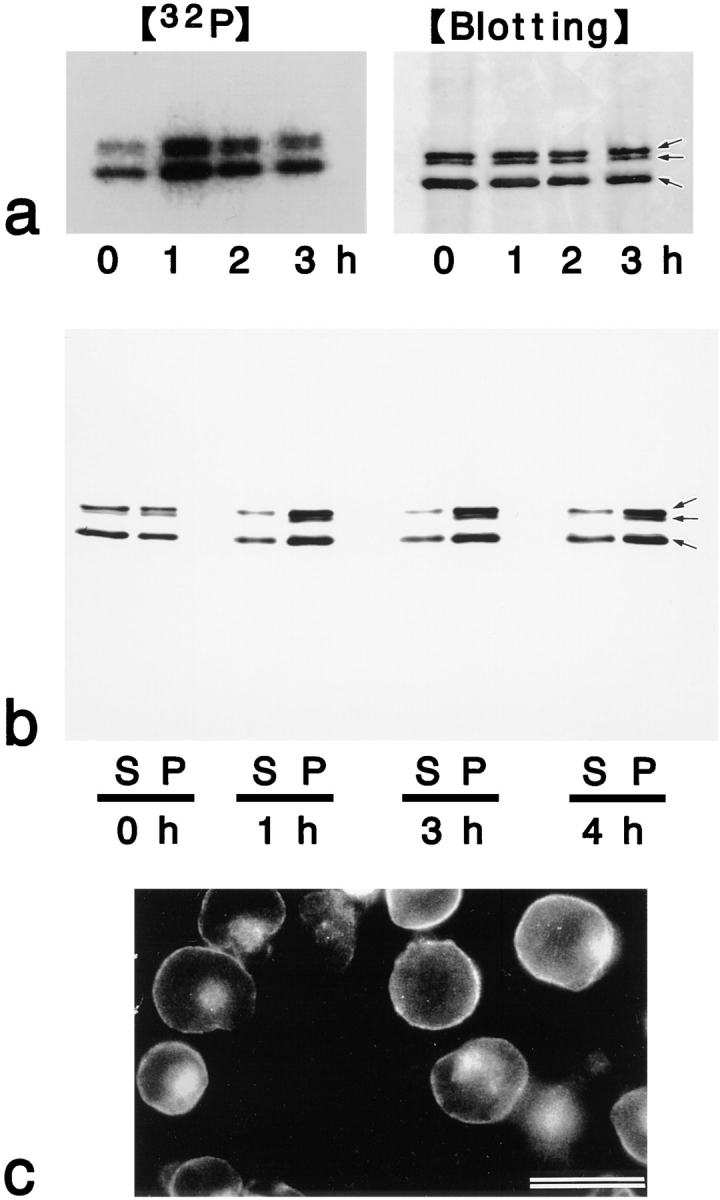
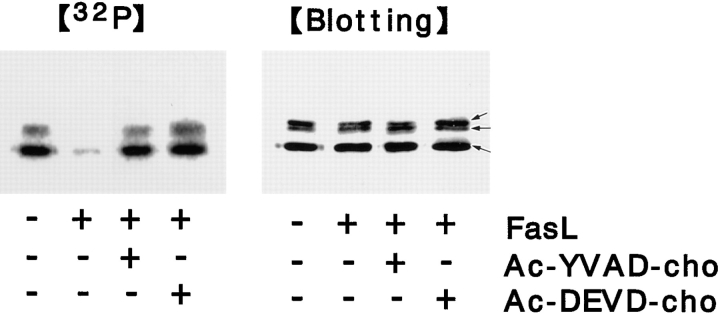
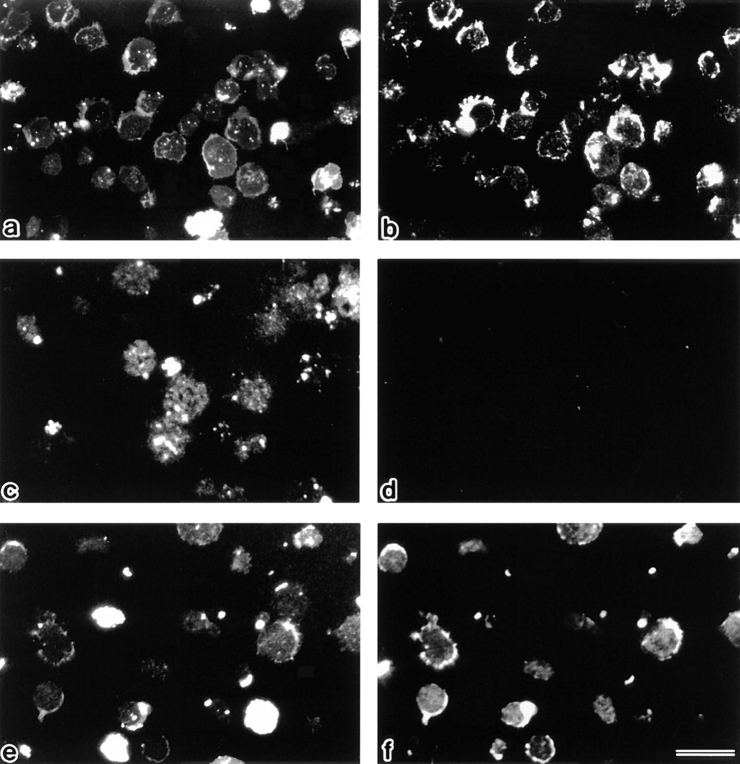
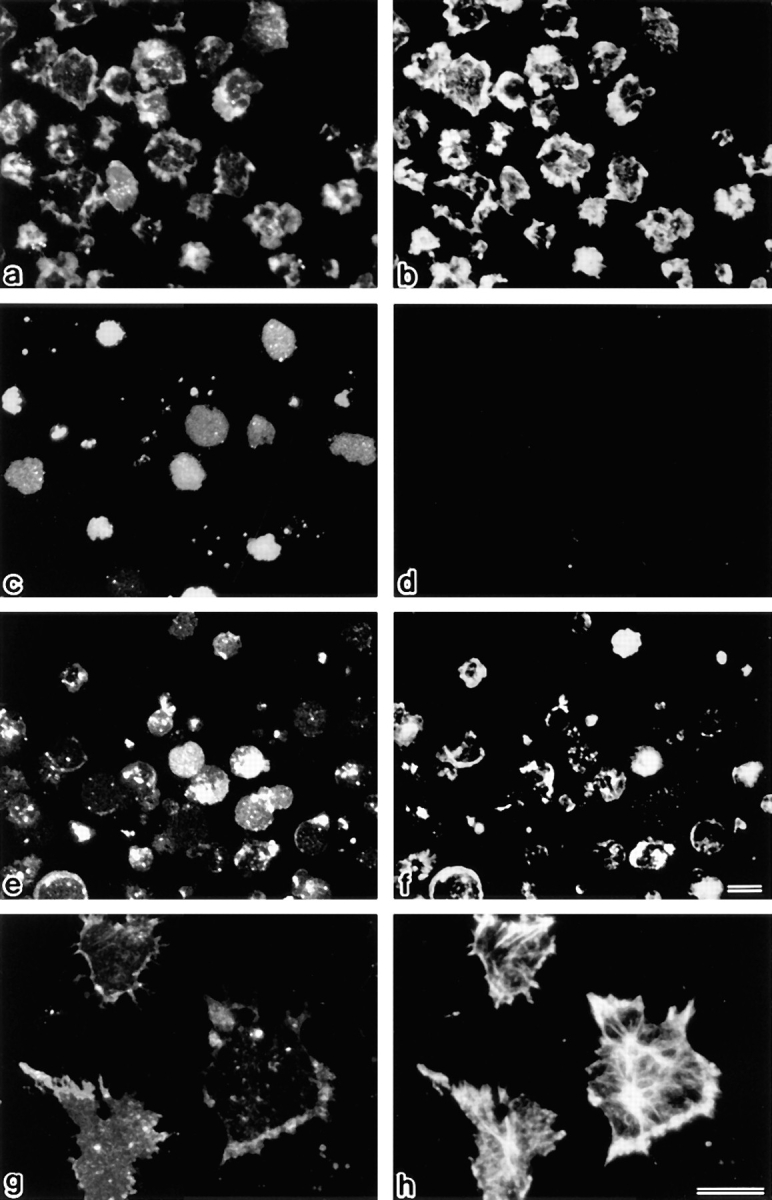
Breakdown of microvilli is a common early event in various types of apoptosis, but its molecular mechanism and implications remain unclear. ERM (ezrin/radixin/moesin) proteins are ubiquitously expressed microvillar proteins that are activated in the cytoplasm, translocate to the plasma membrane, and function as general actin filament/plasma membrane cross-linkers to form microvilli. Immunofluorescence microscopic and biochemical analyses revealed that, at the early phase of Fas ligand (FasL)-induced apoptosis in L cells expressing Fas (LHF), ERM proteins translocate from the plasma membranes of microvilli to the cytoplasm concomitant with dephosphorylation. When the FasL-induced dephosphorylation of ERM proteins was suppressed by calyculin A, a serine/threonine protein phosphatase inhibitor, the cytoplasmic translocation of ERM proteins was blocked. The interleukin-1beta-converting enzyme (ICE) protease inhibitors suppressed the dephosphorylation as well as the cytoplasmic translocation of ERM proteins. These findings indicate that during FasL-induced apoptosis, the ICE protease cascade was first activated, and then ERM proteins were dephosphorylated followed by their cytoplasmic translocation, i.e., microvillar breakdown. Next, to examine the subsequent events in microvillar breakdown, we prepared DiO-labeled single-layered plasma membranes with the cytoplasmic surface freely exposed from FasL-treated or nontreated LHF cells. On single-layered plasma membranes from nontreated cells, ERM proteins and actin filaments were densely detected, whereas those from FasL-treated cells were free from ERM proteins or actin filaments. We thus concluded that the cytoplasmic translocation of ERM proteins is responsible for the microvillar breakdown at an early phase of apoptosis and that the depletion of ERM proteins from plasma membranes results in the gross dissociation of actin-based cytoskeleton from plasma membranes. The physiological relevance of this ERM protein-based microvillar breakdown in apoptosis will be discussed.
Figures
References
-
- Alnemri ES, Livingston DJ, Nicholoson DW, Salvensen G, Thornberry NA, Wong WW, Yuan J. Human ICE-CED-3 protease nomenclature. Cell. 1996;87:171. - PubMed
-
- Arpin M, Algrain M, Louvard D. Membrane-actin microfilament connections: an incresing diversity of players related to band 4.1. Curr Opin Cell Biol. 1994;6:136–141. - PubMed
-
- Boyle WJ, van der Geer P, Hunter T. Phosphopeptide mapping and phosphoamino acid analysis by two-dimensional separation on thin-layer cellulose plates. Methods Enzymol. 1991;201:110–148. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
