

Dismantling cell-cell contacts during apoptosis is coupled to a caspase-dependent proteolytic cleavage of beta-catenin
- PMID: 9348292
- PMCID: PMC2141701
- DOI: 10.1083/jcb.139.3.759
Dismantling cell-cell contacts during apoptosis is coupled to a caspase-dependent proteolytic cleavage of beta-catenin
Abstract
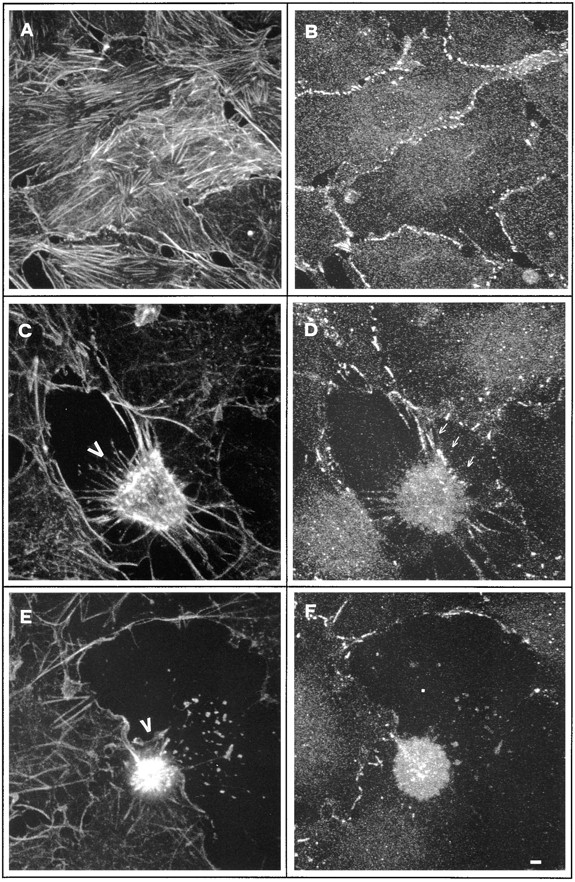
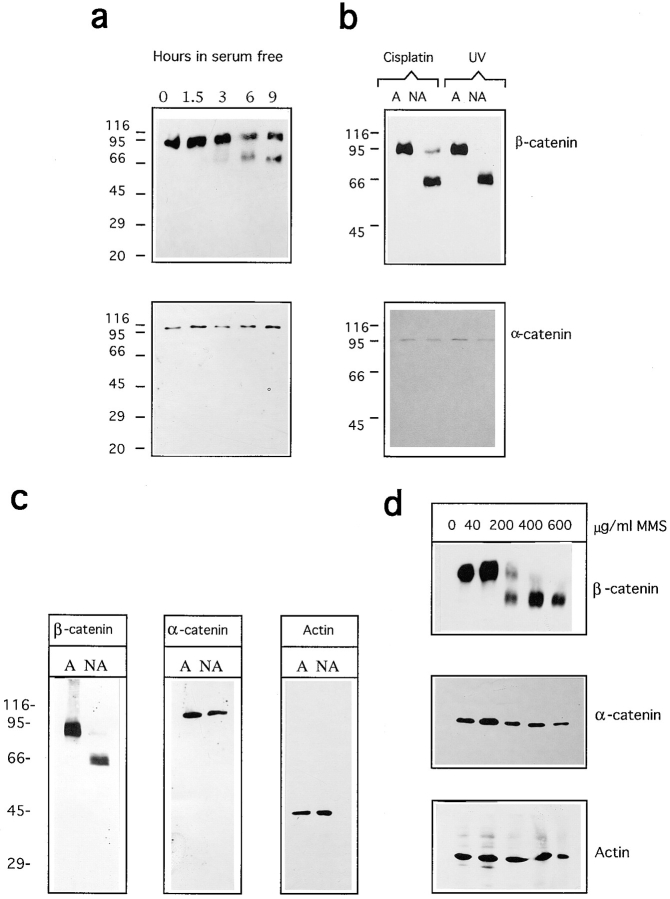
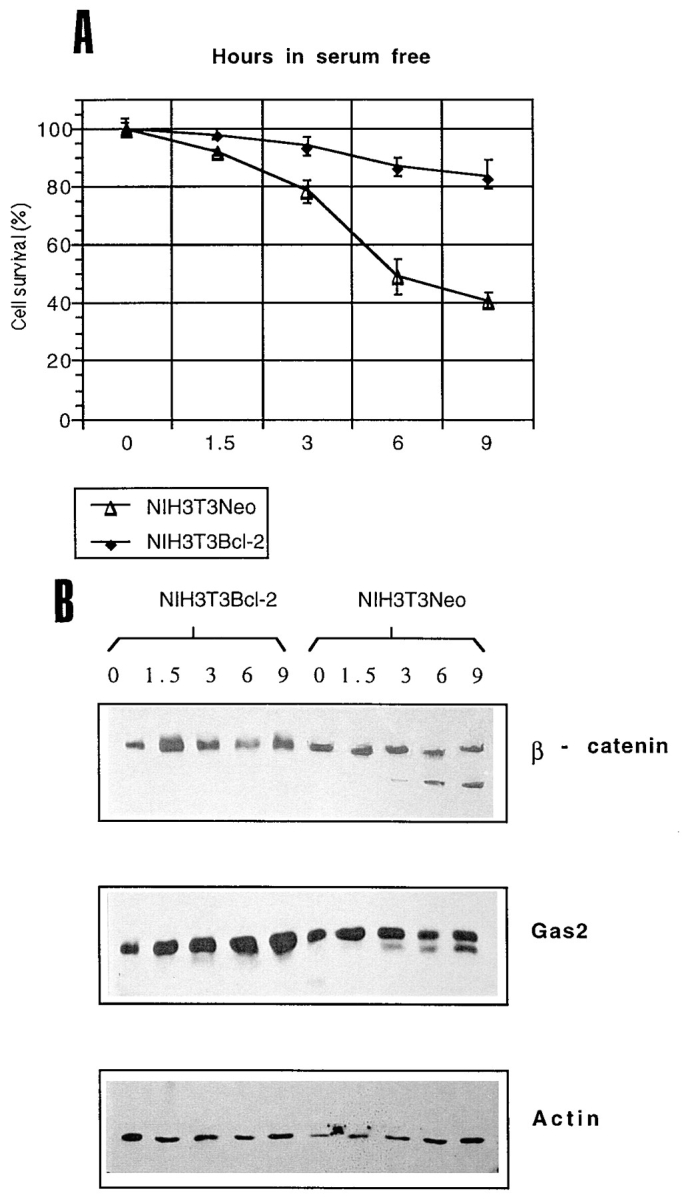
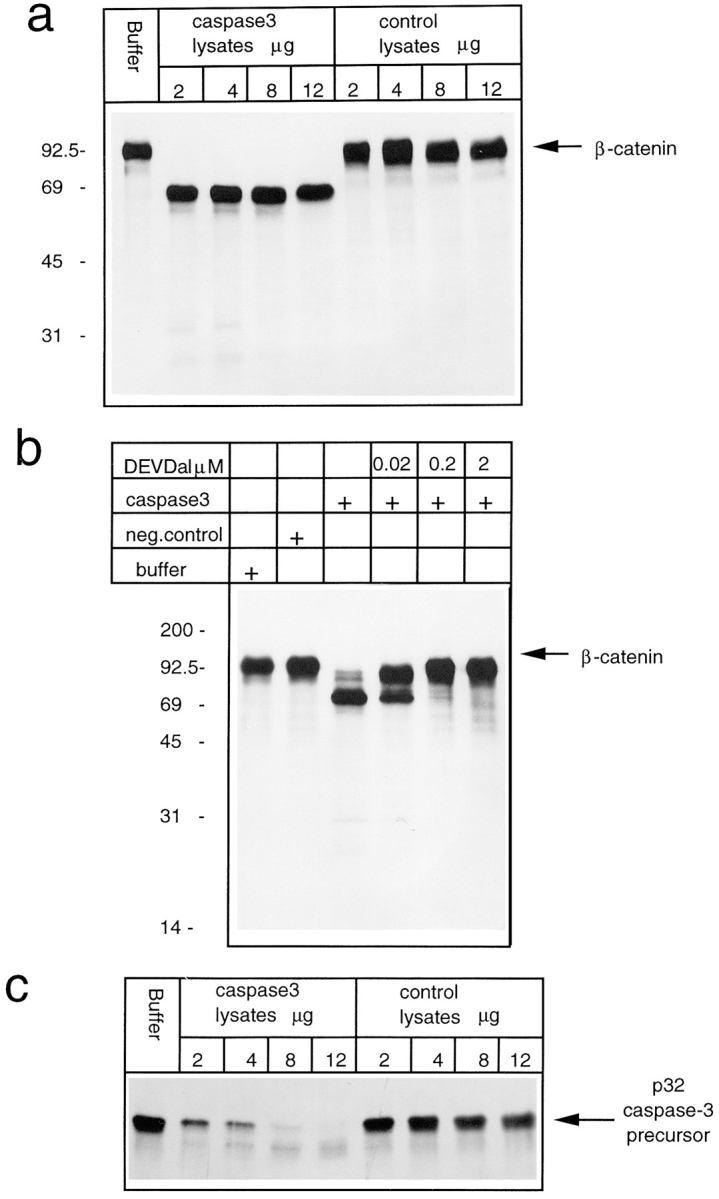
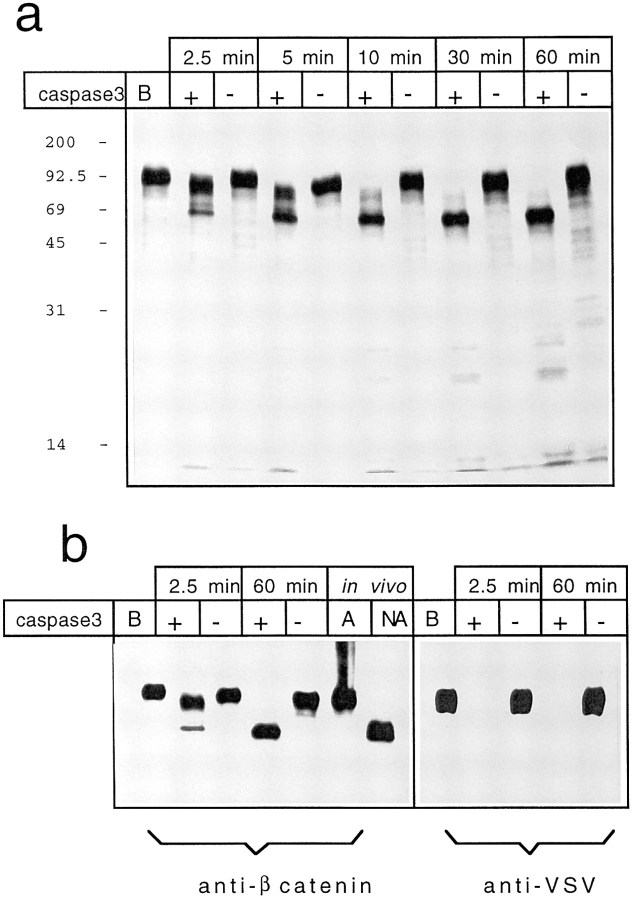
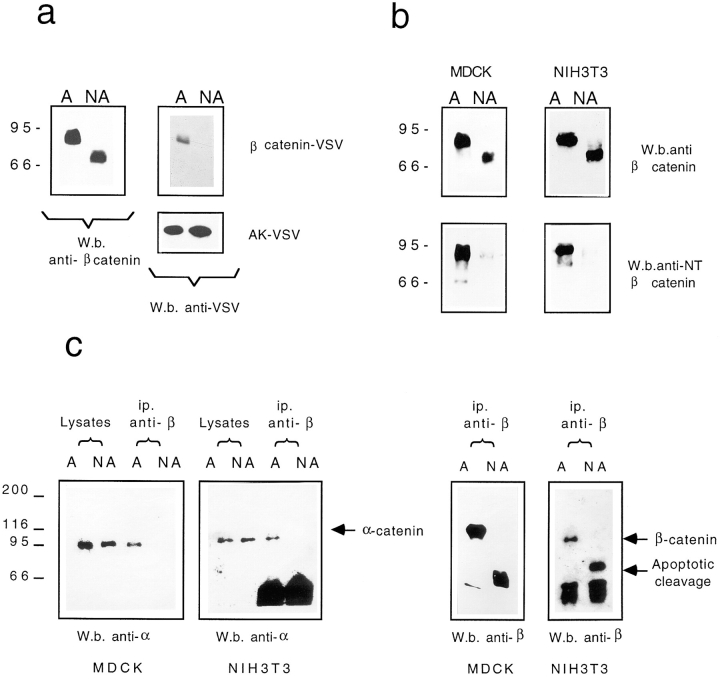
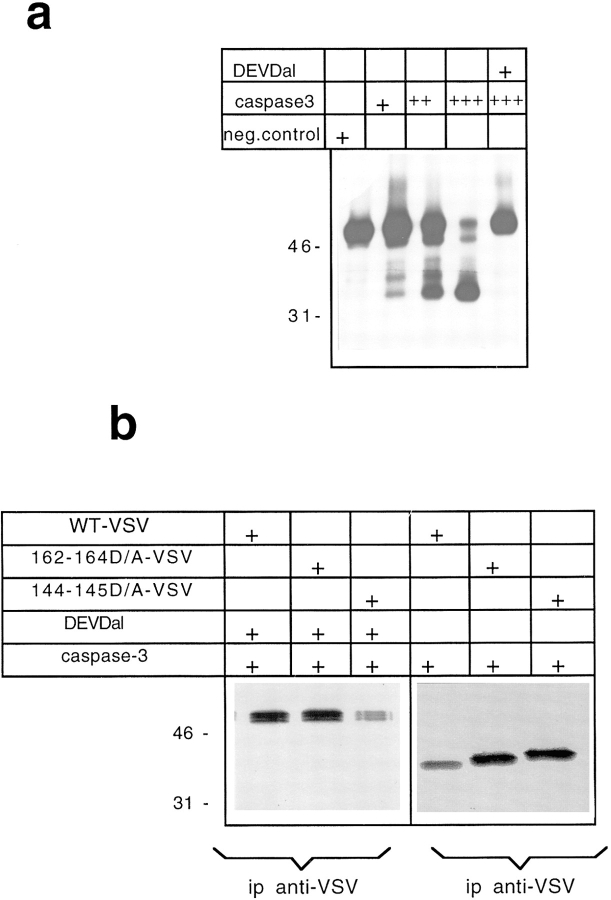
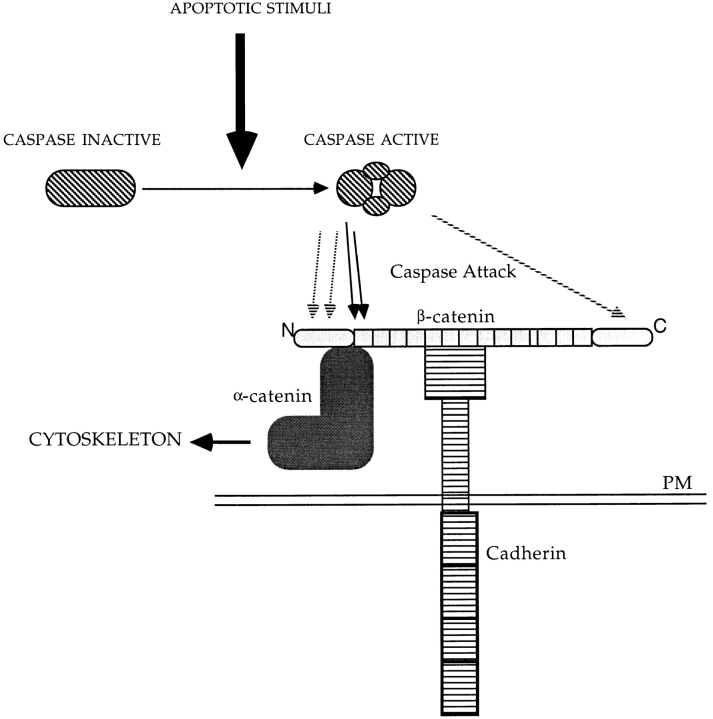
Cell death by apoptosis is a tightly regulated process that requires coordinated modification in cellular architecture. The caspase protease family has been shown to play a key role in apoptosis. Here we report that specific and ordered changes in the actin cytoskeleton take place during apoptosis. In this context, we have dissected one of the first hallmarks in cell death, represented by the severing of contacts among neighboring cells. More specifically, we provide demonstration for the mechanism that could contribute to the disassembly of cytoskeletal organization at cell-cell adhesion. In fact, beta-catenin, a known regulator of cell-cell adhesion, is proteolytically processed in different cell types after induction of apoptosis. Caspase-3 (cpp32/apopain/yama) cleaves in vitro translated beta-catenin into a form which is similar in size to that observed in cells undergoing apoptosis. beta-Catenin cleavage, during apoptosis in vivo and after caspase-3 treatment in vitro, removes the amino- and carboxy-terminal regions of the protein. The resulting beta-catenin product is unable to bind alpha-catenin that is responsible for actin filament binding and organization. This evidence indicates that connection with actin filaments organized at cell-cell contacts could be dismantled during apoptosis. Our observations suggest that caspases orchestrate the specific and sequential changes in the actin cytoskeleton occurring during cell death via cleavage of different regulators of the microfilament system.
Figures



References
-
- Aberle H, Butz S, Stappert J, Weissig H, Kemler R, Hoschuetzky H. Assembling of the cadherin catenin complex in vitro with recombinant proteins. J Cell Sci. 1994;107:3655–3663. - PubMed
-
- Alnemri ES, Livingston DJ, Nicholson DW, Salvesen G, Thornberry NA, Wong WW, Yuan J. Human ICE/CED3 protease nomenclature. Cell. 1996;87:171. - PubMed
-
- Behrens J, von Kries JP, Kuhl M, Bruhn L, Wedlich D, Grosschedl R, Birchmeier W. Functional interaction of β-catenin with the transcription factor LEF-1 . Nature (Lond) 1996;382:225–230. - PubMed
-
- Beidler D, Tewari M, Friesen P, Poirier GG, Dixit V. The baculovirus p35 protein inhibits Fas- and tumor necrosis factor-induced apoptosis. J Biol Chem. 1995;270:16526–16528. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
