

Correlation between charge movement and ionic current during slow inactivation in Shaker K+ channels
- PMID: 9348329
- PMCID: PMC2229383
- DOI: 10.1085/jgp.110.5.579
Correlation between charge movement and ionic current during slow inactivation in Shaker K+ channels
Abstract
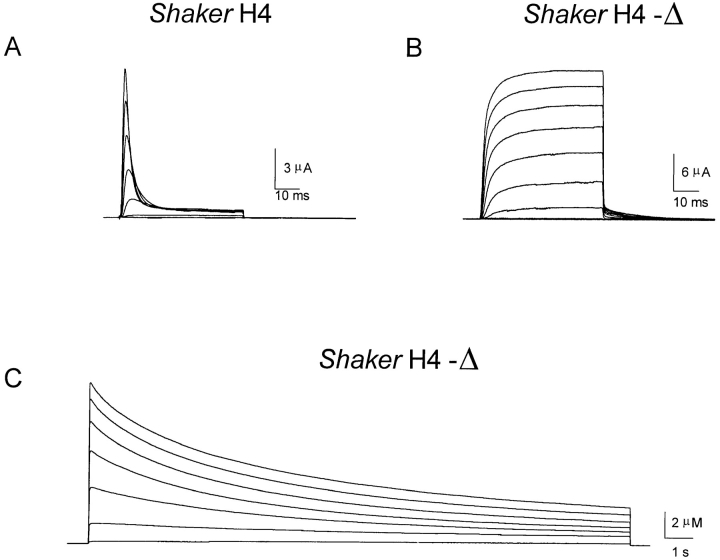
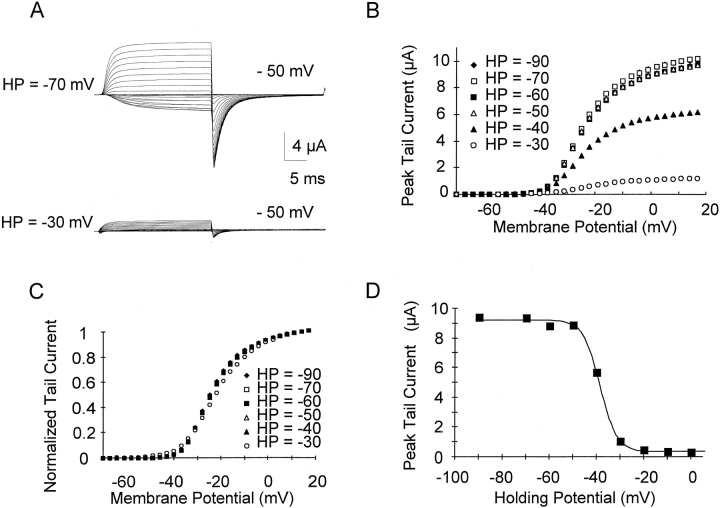
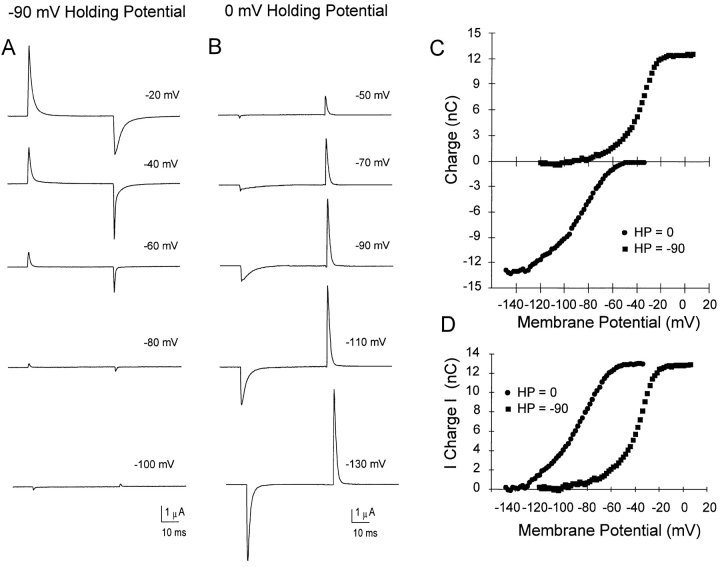
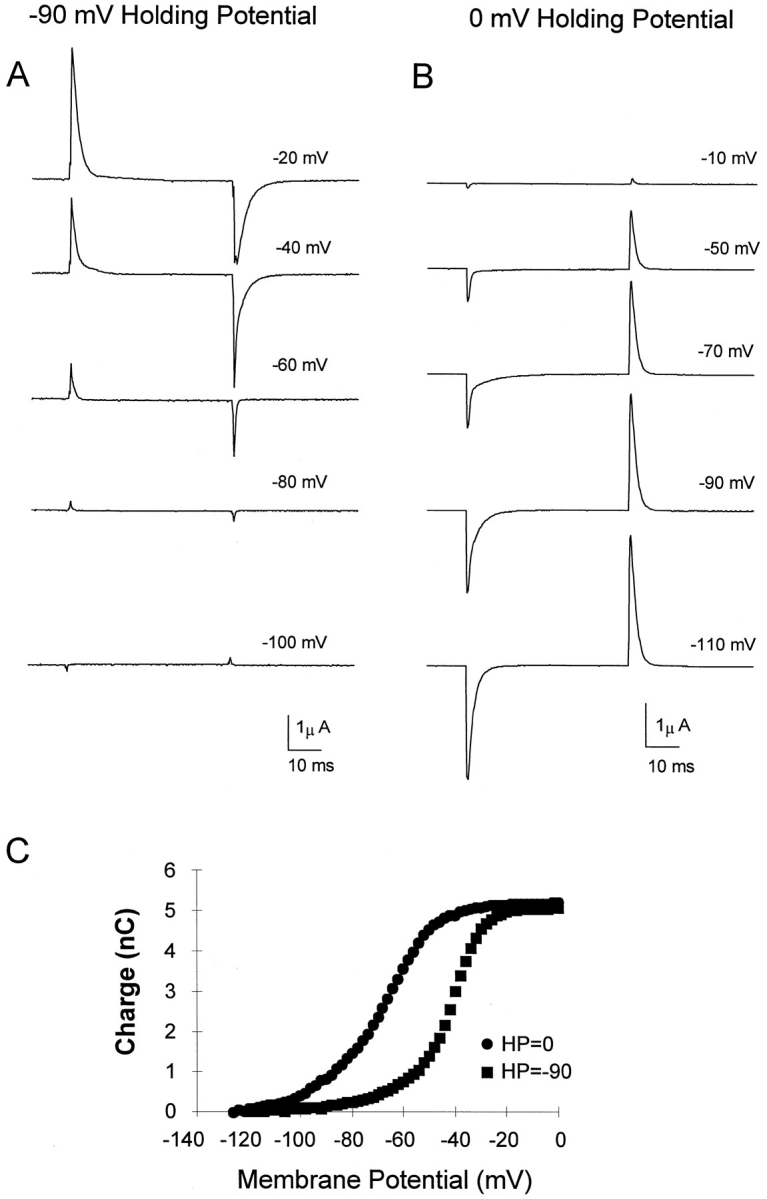
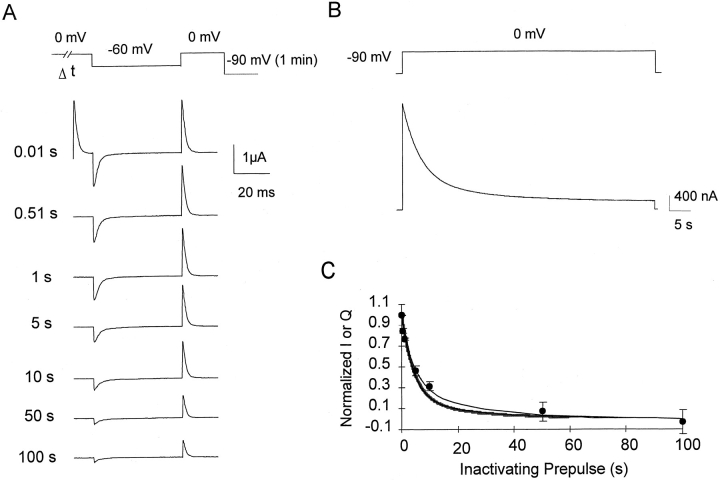
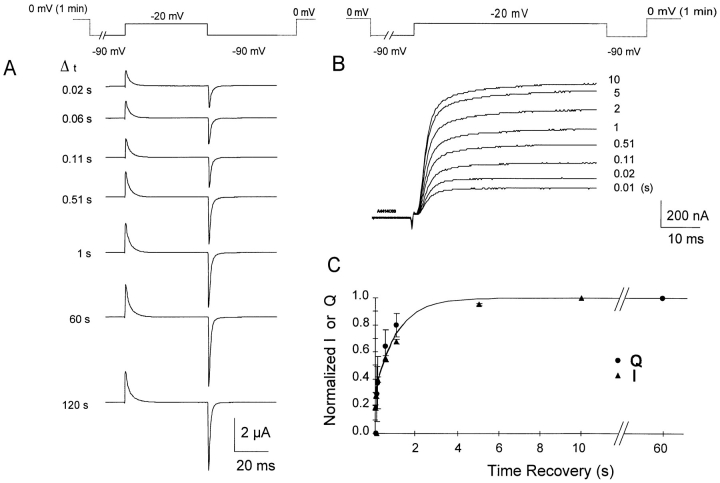
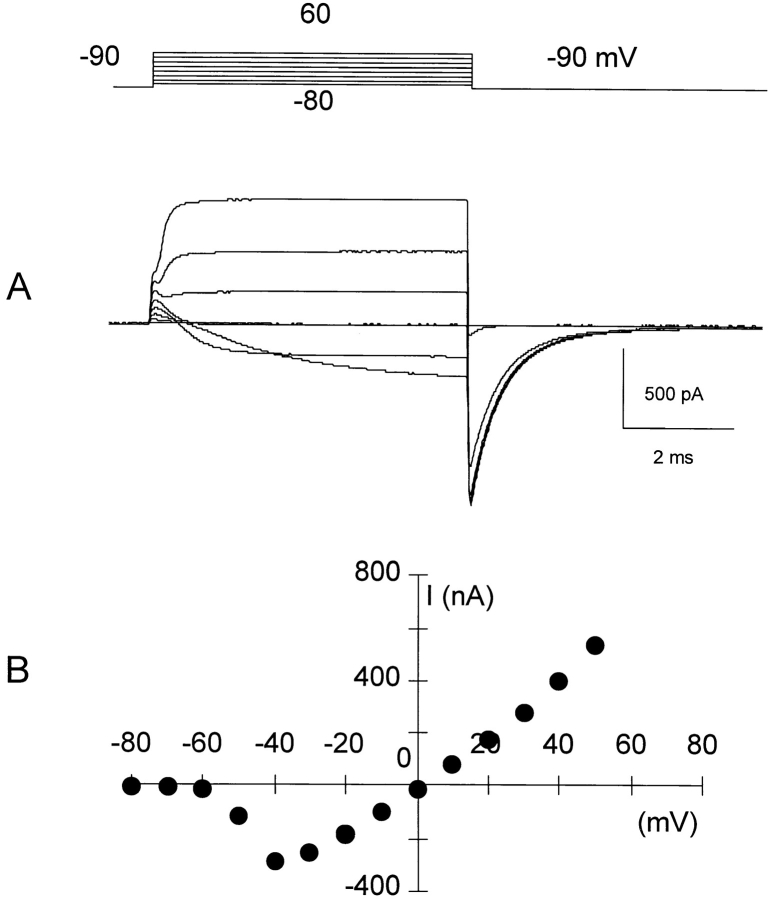
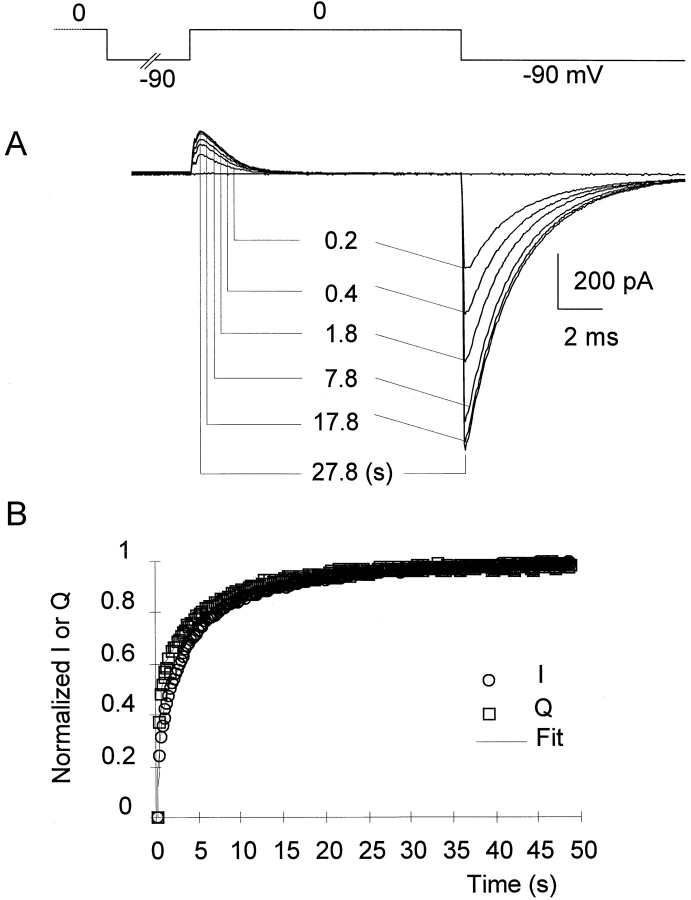
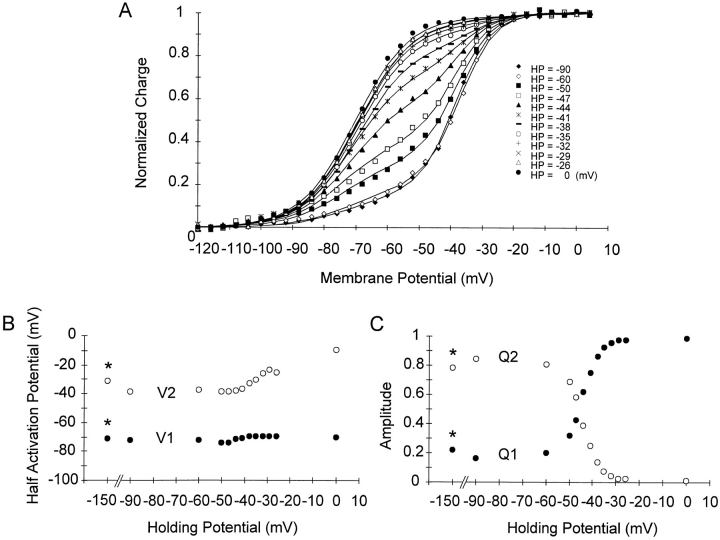
Prolonged depolarization induces a slow inactivation process in some K+ channels. We have studied ionic and gating currents during long depolarizations in the mutant Shaker H4-Delta(6-46) K+ channel and in the nonconducting mutant (Shaker H4-Delta(6-46)-W434F). These channels lack the amino terminus that confers the fast (N-type) inactivation (Hoshi, T., W.N. Zagotta, and R.W. Aldrich. 1991. Neuron. 7:547-556). Channels were expressed in oocytes and currents were measured with the cut-open-oocyte and patch-clamp techniques. In both clones, the curves describing the voltage dependence of the charge movement were shifted toward more negative potentials when the holding potential was maintained at depolarized potentials. The evidences that this new voltage dependence of the charge movement in the depolarized condition is associated with the process of slow inactivation are the following: (a) the installation of both the slow inactivation of the ionic current and the inactivation of the charge in response to a sustained 1-min depolarization to 0 mV followed the same time course; and (b) the recovery from inactivation of both ionic and gating currents (induced by repolarizations to -90 mV after a 1-min inactivating pulse at 0 mV) also followed a similar time course. Although prolonged depolarizations induce inactivation of the majority of the channels, a small fraction remains non-slow inactivated. The voltage dependence of this fraction of channels remained unaltered, suggesting that their activation pathway was unmodified by prolonged depolarization. The data could be fitted to a sequential model for Shaker K+ channels (Bezanilla, F., E. Perozo, and E. Stefani. 1994. Biophys. J. 66:1011-1021), with the addition of a series of parallel nonconducting (inactivated) states that become populated during prolonged depolarization. The data suggest that prolonged depolarization modifies the conformation of the voltage sensor and that this change can be associated with the process of slow inactivation.
Figures

References
-
- Bezanilla F, Perozo E, Papazian DM, Stefani E. Molecular bases of gating charge immobilization in Shakerpotassium channel. Science (Wash DC) 1991;254:679–683. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
