

Peroxide modulation of slow onset potentiation in rat hippocampus
- PMID: 9348338
- PMCID: PMC6573091
- DOI: 10.1523/JNEUROSCI.17-22-08695.1997
Peroxide modulation of slow onset potentiation in rat hippocampus
Abstract
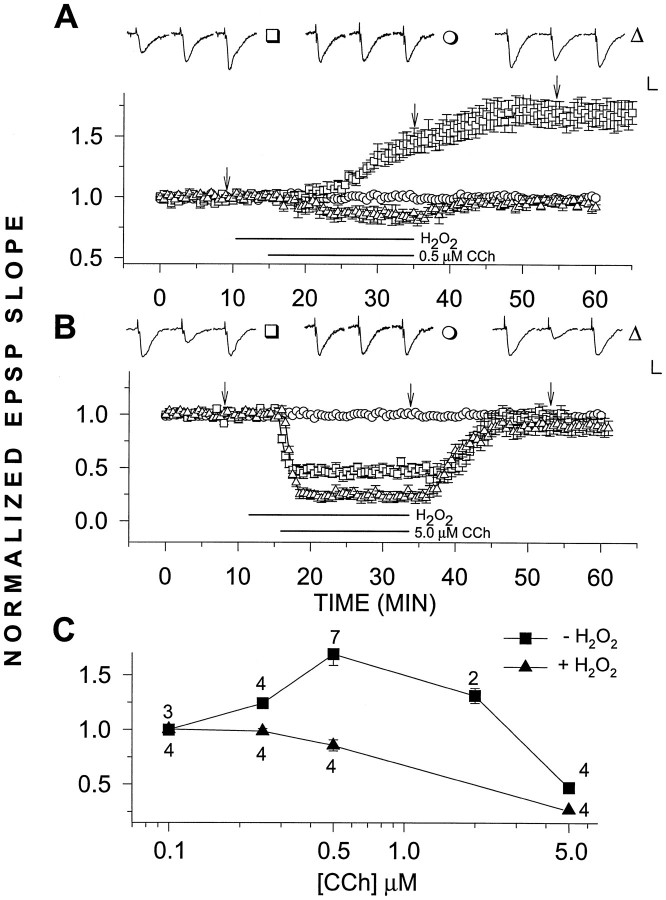
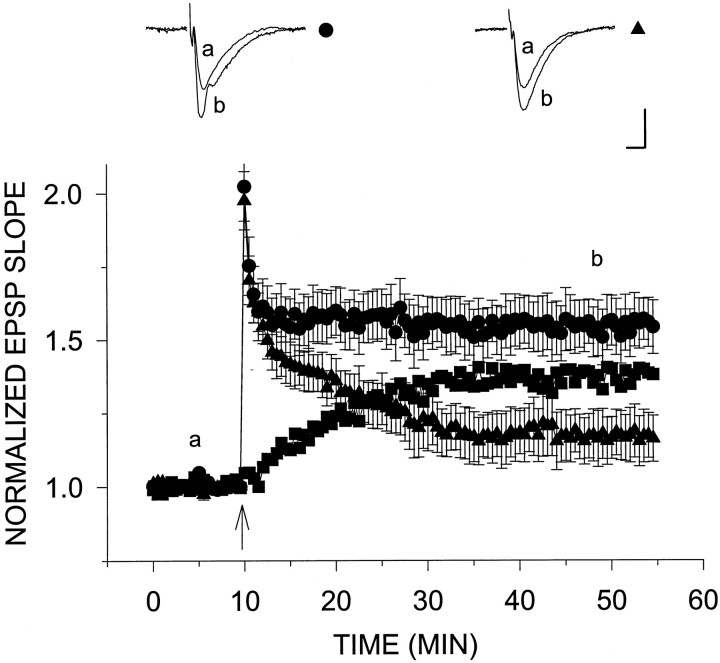
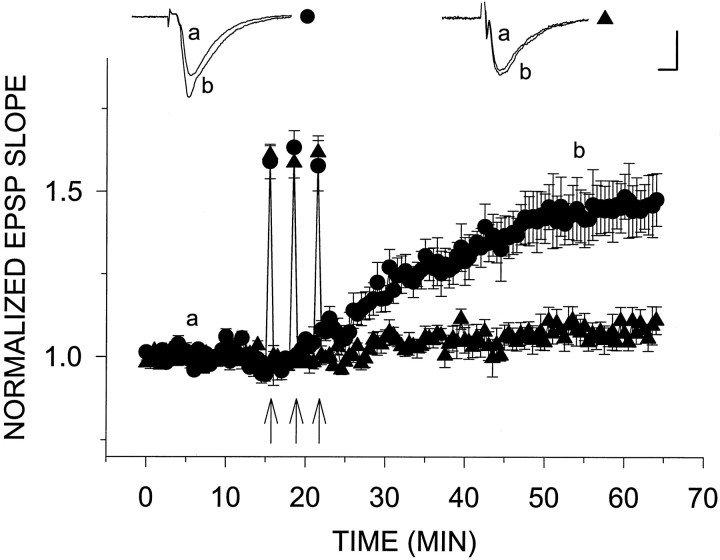
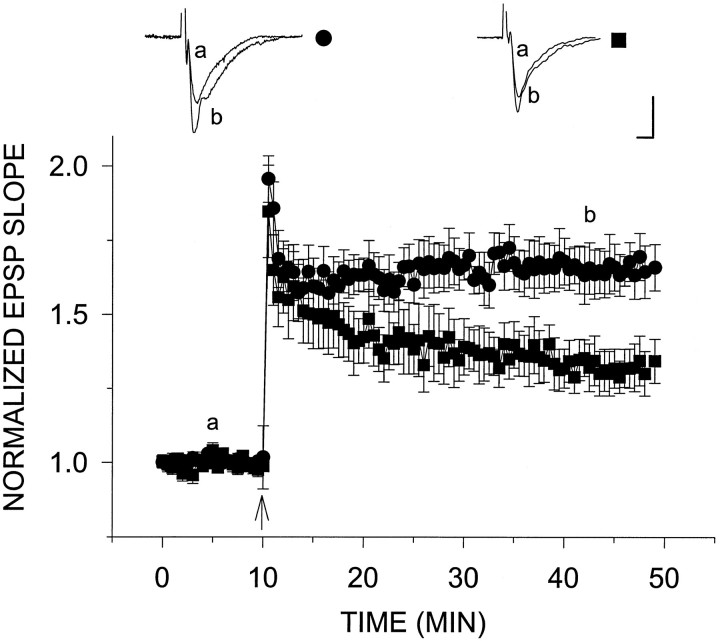
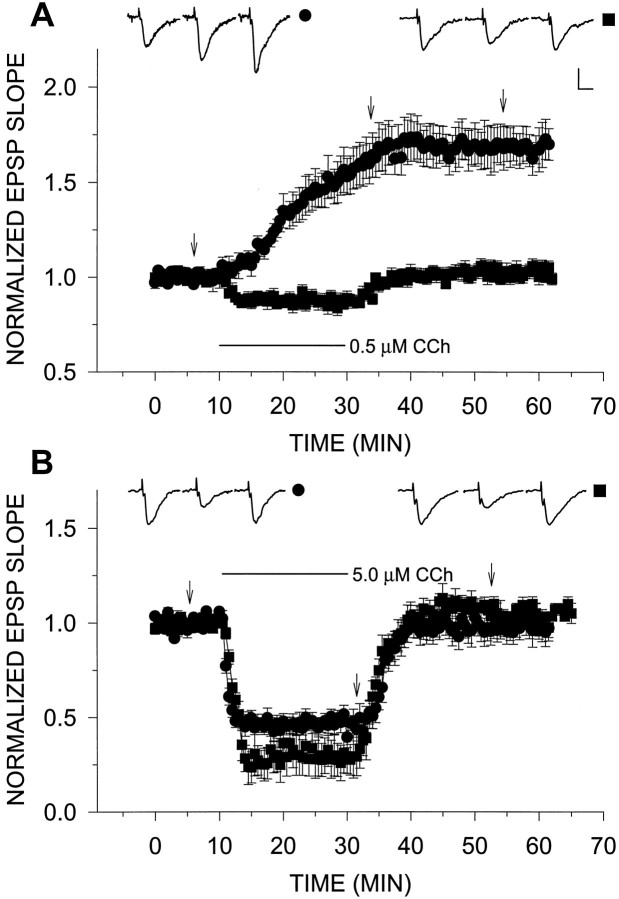
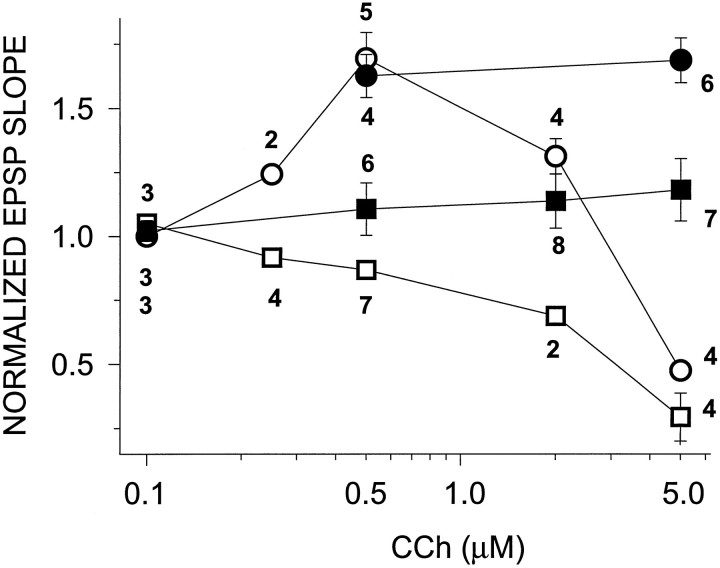
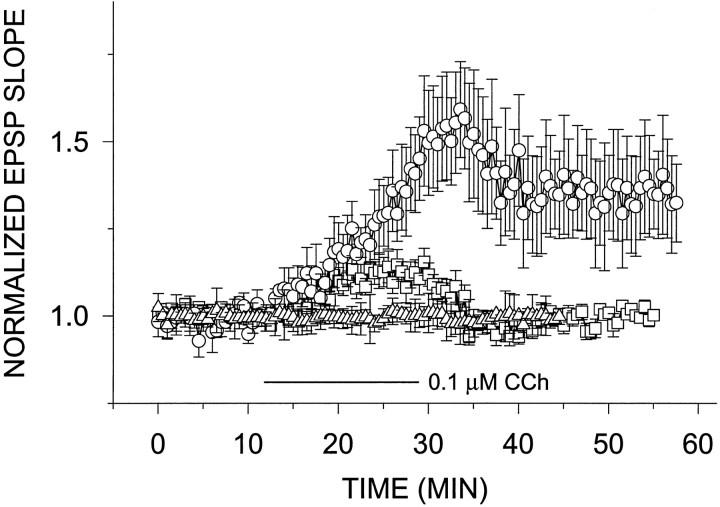
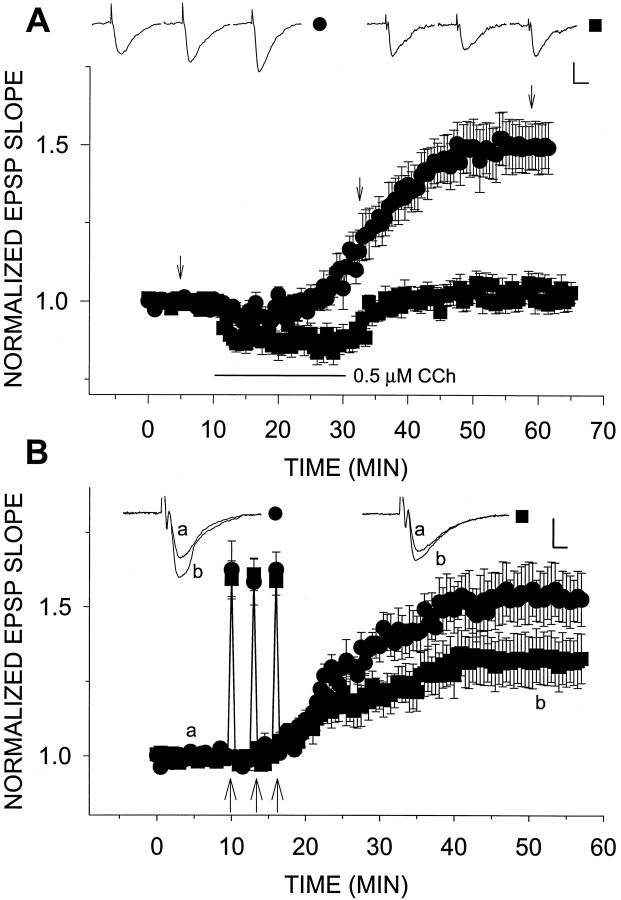
Exposure of rat hippocampal slices to low concentrations of the muscarinic agonist carbachol (CCh) has been shown to produce a slow onset long-term potentiation (LTP) of reactivity to afferent stimulation in CA1 neurons. Although this potentiation shares a number of properties with tetanic LTP, muscarinic LTP (LTPm) is independent of activation of the NMDA receptor. We now demonstrate that low levels of hydrogen peroxide (H2O2) cause hippocampal slices to lose the ability to express LTPm. This powerful effect of H2O2 is selective in that it does not affect the reactivity of hippocampal neurons to higher concentrations of CCh. In fact, H2O2 also blocks induction of a slow onset, non-NMDA-dependent tetanic LTP (NN-LTP). The functional relevance of this action of H2O2 is exemplified by the fact that the hippocampus of aged rats, which produces higher levels of endogenous H2O2 than that of young rats, lacks LTPm and expresses a markedly reduced NN-LTP. In aged rats, the lack of LTPm contrasts with an apparently normal muscarinic suppression of the EPSP slope induced by higher concentrations of CCh. When hippocampal slices from aged animals are treated with catalase, an enzyme that breaks down H2O2, LTPm is restored, and NN-LTP is enhanced. Thus, our study proposes a unique and novel age-dependent peroxide regulation of LTPm in the brain and provides a link between the cholinergic system, aging, and memory functions.
Figures
References
-
- Aniksztejn L, Ben-Ari Y. Novel form of long-term potentiation produced by a K+ channel blocker in the hippocampus. Nature. 1991;349:67–69. - PubMed
-
- Auerbach JM, Segal M. A novel cholinergic induction of long-term potentiation in rat hippocampus. J Neurophysiol. 1994;72:2034–2040. - PubMed
-
- Barnes CA. Normal aging: regionally specific changes in hippocampal synaptic transmission. Trends Neurosci. 1994;17:13–18. - PubMed
-
- Bartus RT, Reginald L, Dean RL, Beer B, Lippa AS. The cholinergic hypothesis of geriatric memory dysfunction. Science. 1982;217:408–417. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous